

Molecular de-extinction of ancient antimicrobial peptides enabled by machine learning
- PMID: 37516110
- PMCID: PMC11625410
- DOI: 10.1016/j.chom.2023.07.001
Molecular de-extinction of ancient antimicrobial peptides enabled by machine learning
Abstract
Molecular de-extinction could offer avenues for drug discovery by reintroducing bioactive molecules that are no longer encoded by extant organisms. To prospect for antimicrobial peptides encrypted within extinct and extant human proteins, we introduce the panCleave random forest model for proteome-wide cleavage site prediction. Our model outperformed multiple protease-specific cleavage site classifiers for three modern human caspases, despite its pan-protease design. Antimicrobial activity was observed in vitro for modern and archaic protein fragments identified with panCleave. Lead peptides showed resistance to proteolysis and exhibited variable membrane permeabilization. Additionally, representative modern and archaic protein fragments showed anti-infective efficacy against A. baumannii in both a skin abscess infection model and a preclinical murine thigh infection model. These results suggest that machine-learning-based encrypted peptide prospection can identify stable, nontoxic peptide antibiotics. Moreover, we establish molecular de-extinction through paleoproteome mining as a framework for antibacterial drug discovery.
Keywords: Denisovan; Neanderthal; antibiotic resistance; antibiotics; antimicrobial peptides; drug discovery; hominins; machine learning; mouse models; protein engineering.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests C.F.N. provides consulting services to Invaio Sciences and is a member of the Scientific Advisory Boards of Nowture S.L. and Phare Bio. The de la Fuente Lab has received research funding or in-kind donations from United Therapeutics, Strata Manufacturing PJSC, and Procter & Gamble, none of which were used in support of this work. C.F.N. is on the Advisory Board of Cell Reports Physical Science. An invention disclosure associated with this work has been filed.
Figures
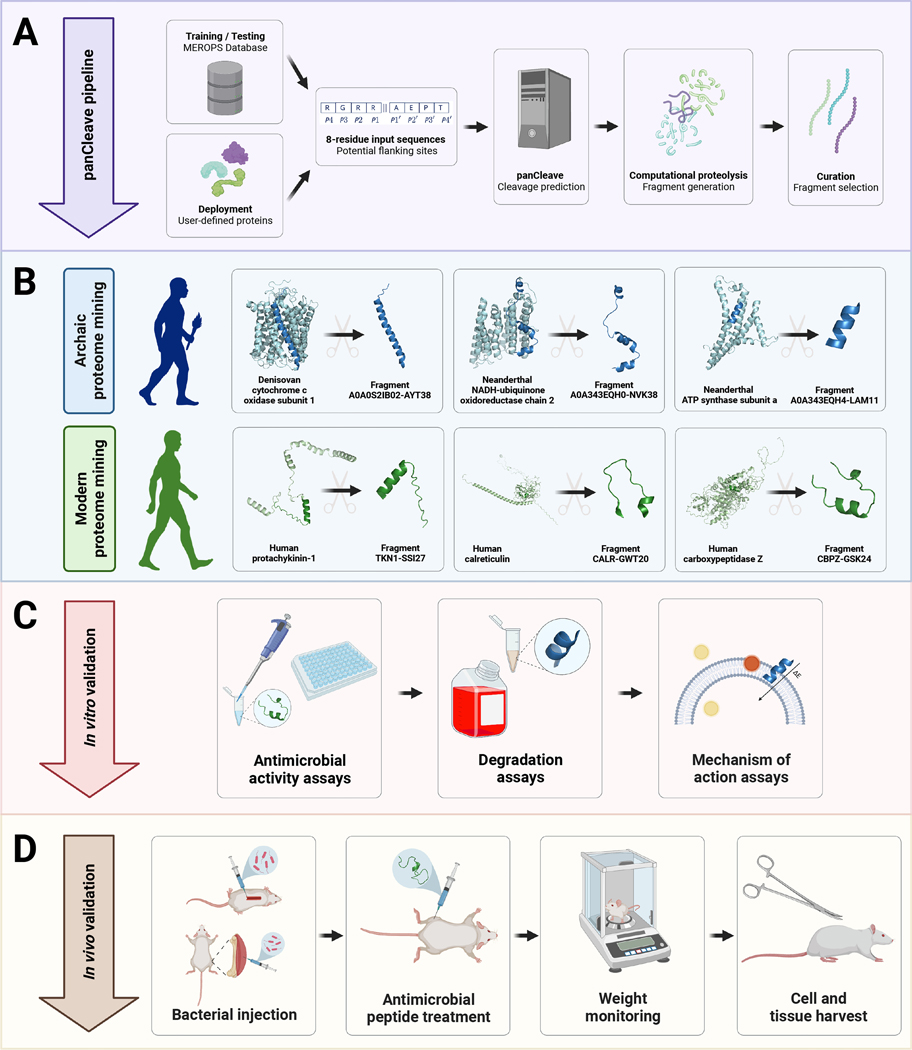
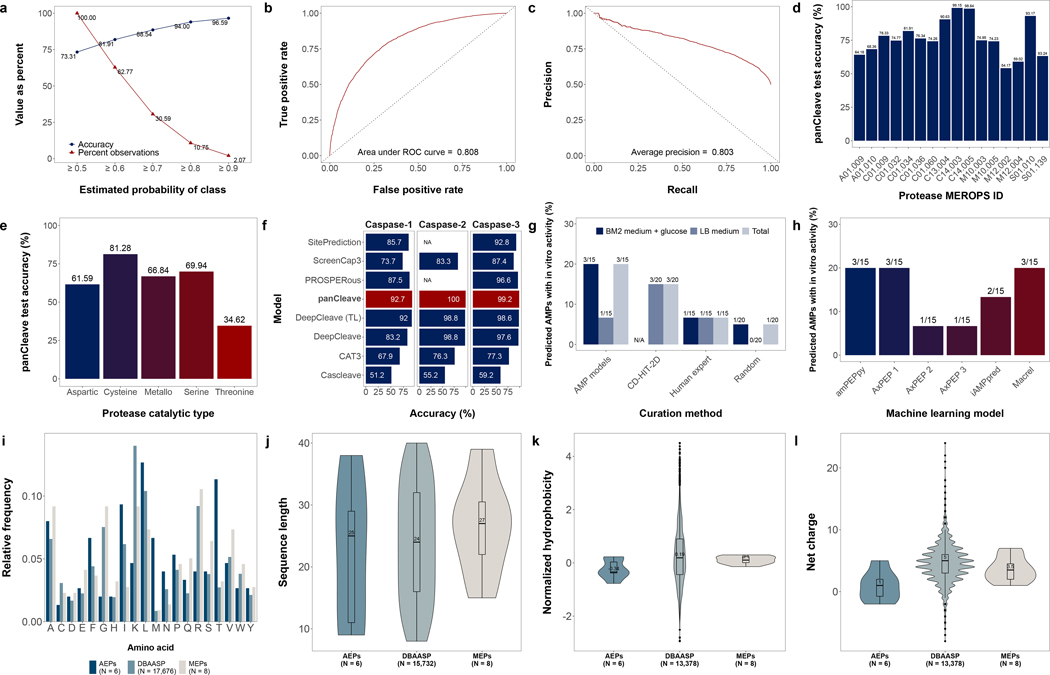
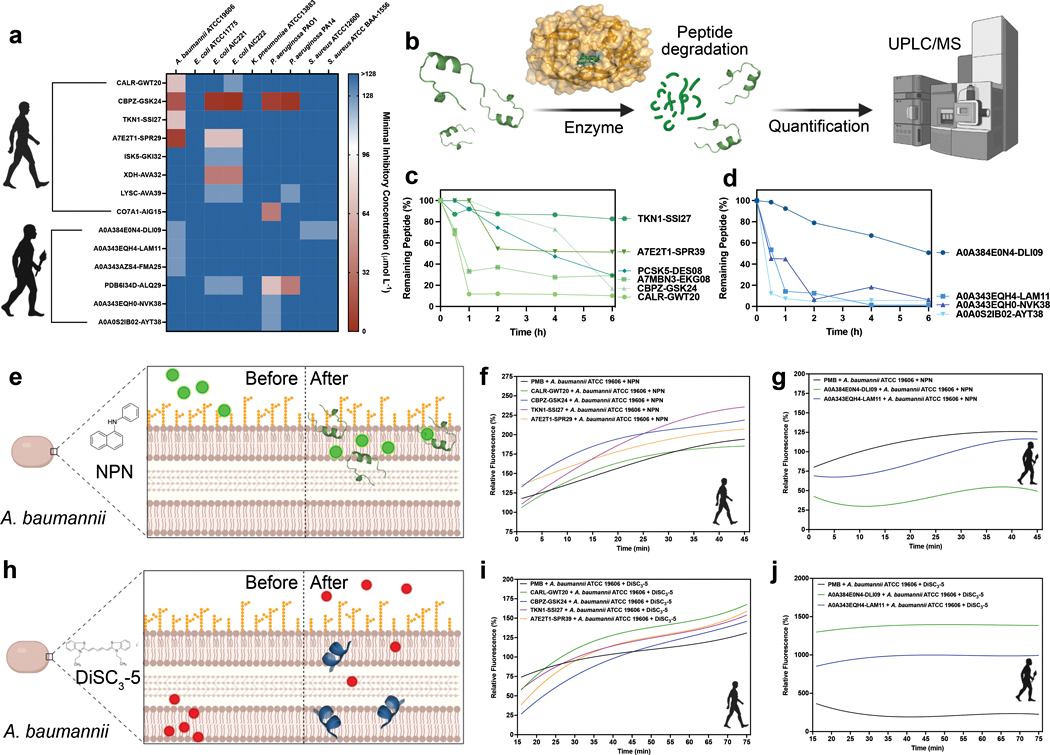
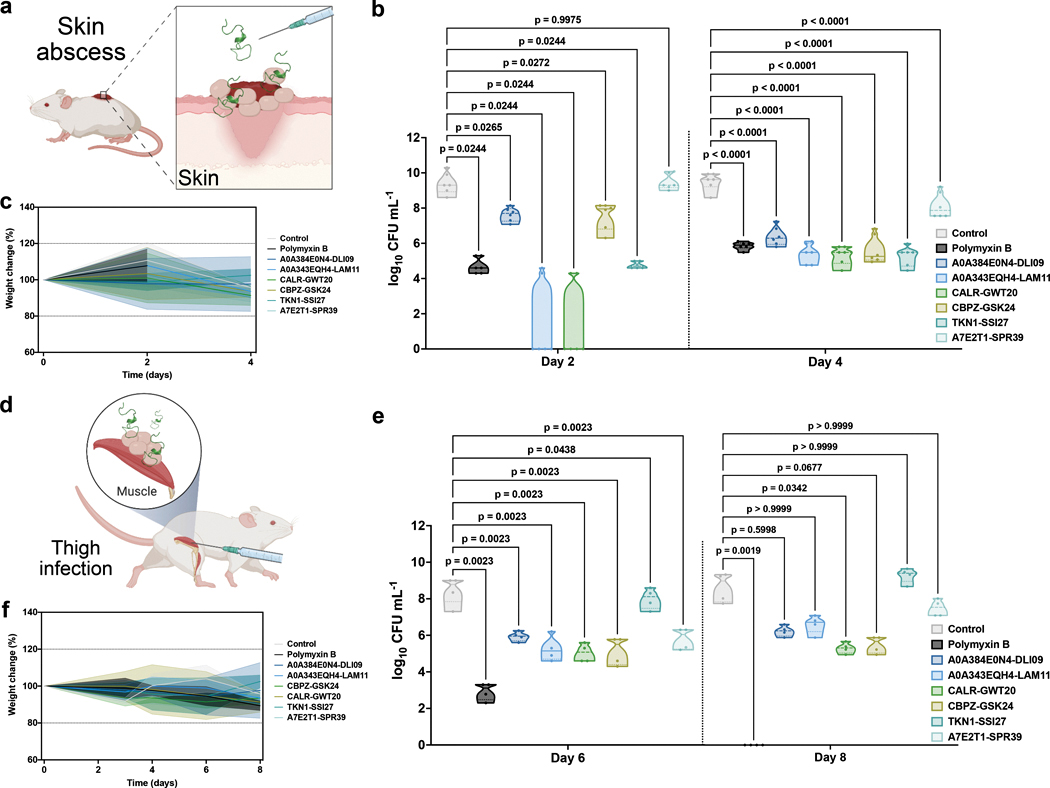
Comment in
-
Mining extinct proteomes for antimicrobial peptides.Nat Biotechnol. 2023 Sep;41(9):1201. doi: 10.1038/s41587-023-01959-6. Nat Biotechnol. 2023. PMID: 37699979 No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
