

An unconventional TOG domain is required for CLASP localization
- PMID: 37516114
- PMCID: PMC10443533
- DOI: 10.1016/j.cub.2023.07.009
An unconventional TOG domain is required for CLASP localization
Abstract
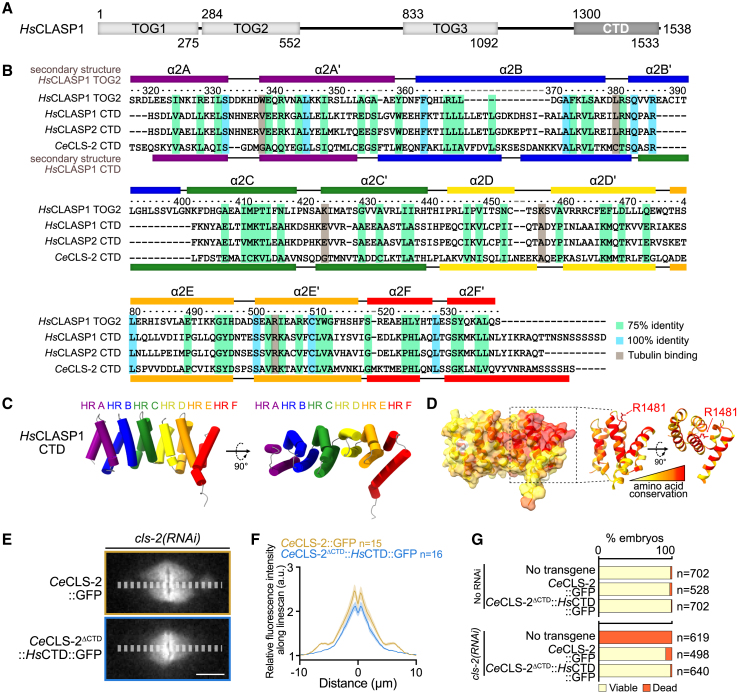
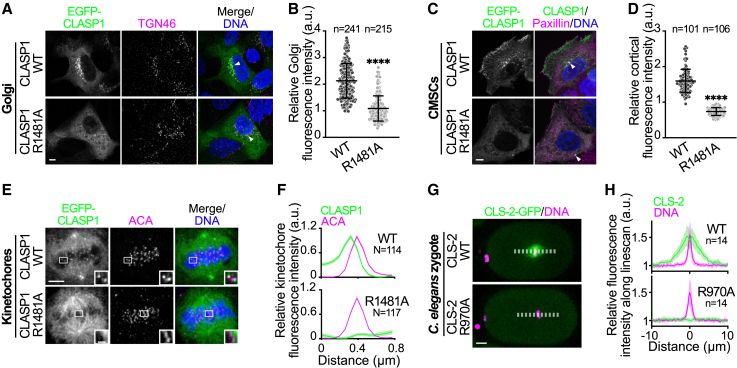
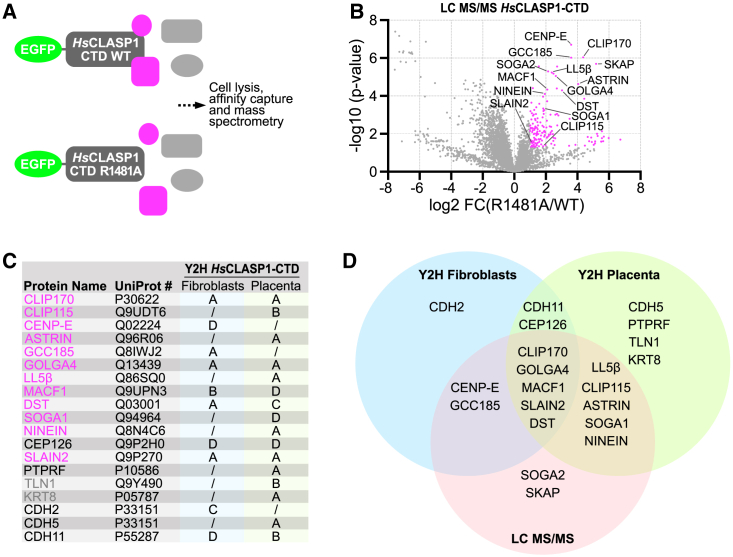
Cytoplasmic linker-associated proteins (CLASPs) form a conserved family of microtubule-associated proteins (MAPs) that maintain microtubules in a growing state by promoting rescue while suppressing catastrophe.1 CLASP function involves an ordered array of tumor overexpressed gene (TOG) domains and binding to multiple protein partners via a conserved C-terminal domain (CTD).2,3 In migrating cells, CLASPs concentrate at the cortex near focal adhesions as part of cortical microtubule stabilization complexes (CMSCs), via binding of their CTD to the focal adhesion protein PHLDB2/LL5β.4,5 Cortical CLASPs also stabilize a subset of microtubules, which stimulate focal adhesion turnover and generate a polarized microtubule network toward the leading edge of migrating cells. CLASPs are also recruited to the trans-Golgi network (TGN) via an interaction between their CTD and the Golgin protein GCC185.6 This allows microtubule growth toward the leading edge of migrating cells, which is required for Golgi organization, polarized intracellular transport, and cell motility.7 In dividing cells, CLASPs are essential at kinetochores for efficient chromosome segregation and anaphase spindle integrity.8,9 Both CENP-E and ASTRIN bind and target CLASPs to kinetochores,10,11 although the CLASP domain required for this interaction is not known. Despite its high evolutionary conservation, the CTD remains structurally uncharacterized. Here, we find that the CTD can be structurally modeled as a TOG domain. We identify a surface-exposed and conserved arginine residue essential for CLASP CTD interaction with partner proteins. Together, our results provide a structural mechanism by which the CLASP CTD directs diverse sub-cellular localizations throughout the cell cycle.
Keywords: CLASP; Golgi; TOG domain; cell division; focal adhesion; microtubules.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Asymmetric CLASP-dependent nucleation of noncentrosomal microtubules at the trans-Golgi network.Dev Cell. 2007 Jun;12(6):917-30. doi: 10.1016/j.devcel.2007.04.002. Dev Cell. 2007. PMID: 17543864 Free PMC article.
-
Prickle1 promotes focal adhesion disassembly in cooperation with the CLASP-LL5β complex in migrating cells.J Cell Sci. 2016 Aug 15;129(16):3115-29. doi: 10.1242/jcs.185439. Epub 2016 Jul 4. J Cell Sci. 2016. PMID: 27378169
-
An isolated CLASP TOG domain suppresses microtubule catastrophe and promotes rescue.Mol Biol Cell. 2018 Jun 1;29(11):1359-1375. doi: 10.1091/mbc.E17-12-0748. Epub 2018 Apr 5. Mol Biol Cell. 2018. PMID: 29851564 Free PMC article.
-
The role of TOG domains in microtubule plus end dynamics.Biochem Soc Trans. 2009 Oct;37(Pt 5):1002-6. doi: 10.1042/BST0371002. Biochem Soc Trans. 2009. PMID: 19754440 Review.
-
Regulation of microtubule dynamics by TOG-domain proteins XMAP215/Dis1 and CLASP.Trends Cell Biol. 2011 Oct;21(10):604-14. doi: 10.1016/j.tcb.2011.06.007. Epub 2011 Jul 23. Trends Cell Biol. 2011. PMID: 21782439 Free PMC article. Review.
Cited by
-
CLASP-mediated competitive binding in protein condensates directs microtubule growth.Nat Commun. 2024 Aug 2;15(1):6509. doi: 10.1038/s41467-024-50863-3. Nat Commun. 2024. PMID: 39095354 Free PMC article.
-
Kinesin-7 CENP-E in tumorigenesis: Chromosome instability, spindle assembly checkpoint, and applications.Front Mol Biosci. 2024 Mar 15;11:1366113. doi: 10.3389/fmolb.2024.1366113. eCollection 2024. Front Mol Biosci. 2024. PMID: 38560520 Free PMC article. Review.
References
-
- Akhmanova A., Hoogenraad C.C., Drabek K., Stepanova T., Dortland B., Verkerk T., Vermeulen W., Burgering B.M., De Zeeuw C.I., Grosveld F., Galjart N. Clasps are CLIP-115 and -170 associating proteins involved in the regional regulation of microtubule dynamics in motile fibroblasts. Cell. 2001;104:923–935. - PubMed
-
- Lansbergen G., Grigoriev I., Mimori-Kiyosue Y., Ohtsuka T., Higa S., Kitajima I., Demmers J., Galjart N., Houtsmuller A.B., Grosveld F., Akhmanova A. CLASPs attach microtubule plus ends to the cell cortex through a complex with LL5beta. Dev. Cell. 2006;11:21–32. doi: 10.1016/j.devcel.2006.05.012. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
