

Clade density and the evolution of diversity-dependent diversification
- PMID: 37516766
- PMCID: PMC10387094
- DOI: 10.1038/s41467-023-39629-5
Clade density and the evolution of diversity-dependent diversification
Abstract
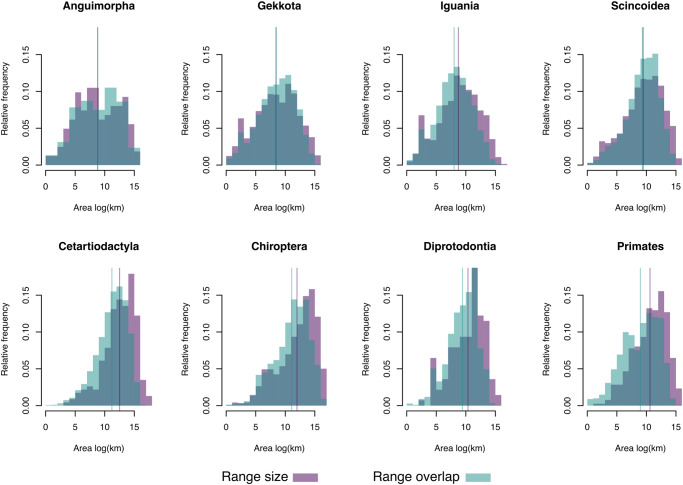
The assumption of an ecological limit to the number of species in a given region is frequently invoked in evolutionary studies, yet its empirical basis is remarkably meager. We explore this assumption by integrating data on geographical distributions and phylogenetic relationships of nearly six thousand terrestrial vertebrate species. In particular, we test whether sympatry with closely-related species leads to decreasing speciation rates. We introduce the concept of clade density, which is the sum of the areas of overlap between a given species and other members of its higher taxon, weighted by their phylogenetic distance. Our results showed that, regardless of the chosen taxon and uncertainty in the phylogenetic relationships between the studied species, there is no significant relationship between clade density and speciation rate. We argue that the mechanistic foundation of diversity-dependent diversification is fragile, and that a better understanding of the mechanisms driving regional species pools is sorely needed.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Elton, C. S. The Ecology Of Animals (Wiley, 1950).
-
- Hutchinson GE. Homage to santa rosalia or why are there so many kinds of animals? Am. Nat. 1959;93:145–159. doi: 10.1086/282070. - DOI
-
- Hutchinson GE. The paradox of the plankton. Am. Nat. 1961;95:137–145. doi: 10.1086/282171. - DOI
-
- Macarthur RH. Patterns of species diversity. Biol. Rev. 1965;40:510–533. doi: 10.1111/j.1469-185X.1965.tb00815.x. - DOI
-
- MacArthur R, Levins R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 1967;101:377–385. doi: 10.1086/282505. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
