

Kv12-encoded K+ channels drive the day-night switch in the repetitive firing rates of SCN neurons
- PMID: 37516908
- PMCID: PMC10373311
- DOI: 10.1085/jgp.202213310
Kv12-encoded K+ channels drive the day-night switch in the repetitive firing rates of SCN neurons
Abstract
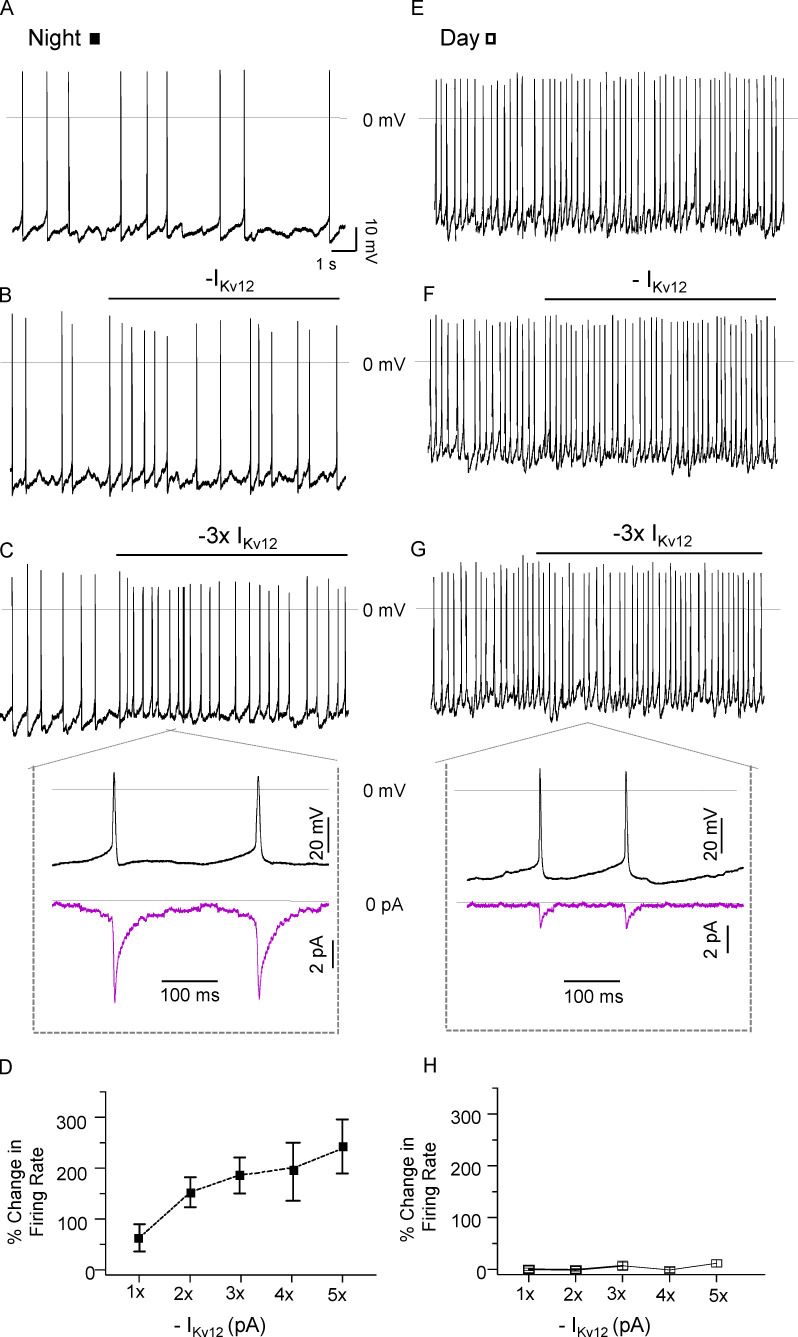
Considerable evidence suggests that day-night rhythms in the functional expression of subthreshold potassium (K+) channels regulate daily oscillations in the spontaneous firing rates of neurons in the suprachiasmatic nucleus (SCN), the master circadian pacemaker in mammals. The K+ conductance(s) driving these daily rhythms in the repetitive firing rates of SCN neurons, however, have not been identified. To test the hypothesis that subthreshold Kv12.1/Kv12.2-encoded K+ channels play a role, we obtained current-clamp recordings from SCN neurons in slices prepared from adult mice harboring targeted disruptions in the Kcnh8 (Kv12.1-/-) or Kcnh3 (Kv12.2-/-) locus. We found that mean nighttime repetitive firing rates were higher in Kv12.1-/- and Kv12.2-/- than in wild type (WT), SCN neurons. In marked contrast, mean daytime repetitive firing rates were similar in Kv12.1-/-, Kv12.2-/-, and WT SCN neurons, and the day-night difference in mean repetitive firing rates, a hallmark feature of WT SCN neurons, was eliminated in Kv12.1-/- and Kv12.2-/- SCN neurons. Similar results were obtained with in vivo shRNA-mediated acute knockdown of Kv12.1 or Kv12.2 in adult SCN neurons. Voltage-clamp experiments revealed that Kv12-encoded current densities in WT SCN neurons are higher at night than during the day. In addition, the pharmacological block of Kv12-encoded currents increased the mean repetitive firing rate of nighttime, but not daytime, in WT SCN neurons. Dynamic clamp-mediated subtraction of modeled Kv12-encoded currents also selectively increased the mean repetitive firing rates of nighttime WT SCN neurons. Despite the elimination of the nighttime decrease in the mean repetitive firing rates of SCN neurons, however, locomotor (wheel-running) activity remained rhythmic in Kv12.1-/-, Kv12.2-/-, and Kv12.1-targeted shRNA-expressing, and Kv12.2-targeted shRNA-expressing animals.
© 2023 Hermanstyne et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures




















Update of
-
Kv12-Encoded K + Channels Drive the Day-Night Switch in the Repetitive Firing Rates of SCN Neurons.bioRxiv [Preprint]. 2023 Feb 2:2023.01.30.526323. doi: 10.1101/2023.01.30.526323. bioRxiv. 2023. Update in: J Gen Physiol. 2023 Sep 4;155(9):e202213310. doi: 10.1085/jgp.202213310. PMID: 36778242 Free PMC article. Updated. Preprint.
Comment in
-
Kv12 channels flick the switch.J Gen Physiol. 2023 Sep 4;155(9):e202313459. doi: 10.1085/jgp.202313459. Epub 2023 Aug 16. J Gen Physiol. 2023. PMID: 37584659 Free PMC article.
Similar articles
-
Effects of NALCN-Encoded Na+ Leak Currents on the Repetitive Firing Properties of SCN Neurons Depend on K+-Driven Rhythmic Changes in Input Resistance.J Neurosci. 2023 Jul 12;43(28):5132-5141. doi: 10.1523/JNEUROSCI.0182-23.2023. Epub 2023 Jun 20. J Neurosci. 2023. PMID: 37339878 Free PMC article.
-
Kv12-Encoded K + Channels Drive the Day-Night Switch in the Repetitive Firing Rates of SCN Neurons.bioRxiv [Preprint]. 2023 Feb 2:2023.01.30.526323. doi: 10.1101/2023.01.30.526323. bioRxiv. 2023. Update in: J Gen Physiol. 2023 Sep 4;155(9):e202213310. doi: 10.1085/jgp.202213310. PMID: 36778242 Free PMC article. Updated. Preprint.
-
Acute Knockdown of Kv4.1 Regulates Repetitive Firing Rates and Clock Gene Expression in the Suprachiasmatic Nucleus and Daily Rhythms in Locomotor Behavior.eNeuro. 2017 May 23;4(3):ENEURO.0377-16.2017. doi: 10.1523/ENEURO.0377-16.2017. eCollection 2017 May-Jun. eNeuro. 2017. PMID: 28560311 Free PMC article.
-
Ion Channels Controlling Circadian Rhythms in Suprachiasmatic Nucleus Excitability.Physiol Rev. 2020 Oct 1;100(4):1415-1454. doi: 10.1152/physrev.00027.2019. Epub 2020 Mar 12. Physiol Rev. 2020. PMID: 32163720 Free PMC article. Review.
-
Linking neural activity and molecular oscillations in the SCN.Nat Rev Neurosci. 2011 Sep 2;12(10):553-69. doi: 10.1038/nrn3086. Nat Rev Neurosci. 2011. PMID: 21886186 Free PMC article. Review.
Cited by
-
Contribution of membrane-associated oscillators to biological timing at different timescales.Front Physiol. 2024 Jan 9;14:1243455. doi: 10.3389/fphys.2023.1243455. eCollection 2023. Front Physiol. 2024. PMID: 38264332 Free PMC article.
-
Effects of NALCN-Encoded Na+ Leak Currents on the Repetitive Firing Properties of SCN Neurons Depend on K+-Driven Rhythmic Changes in Input Resistance.J Neurosci. 2023 Jul 12;43(28):5132-5141. doi: 10.1523/JNEUROSCI.0182-23.2023. Epub 2023 Jun 20. J Neurosci. 2023. PMID: 37339878 Free PMC article.
-
Mechanisms of pacemaking in mammalian neurons.J Physiol. 2024 Sep 20:10.1113/JP284759. doi: 10.1113/JP284759. Online ahead of print. J Physiol. 2024. PMID: 39303139 Review.
-
Inputs and Outputs of the Mammalian Circadian Clock.Biology (Basel). 2023 Mar 28;12(4):508. doi: 10.3390/biology12040508. Biology (Basel). 2023. PMID: 37106709 Free PMC article. Review.
-
KCNH3 Loss-of-Function Variant Associated with Epilepsy and Neurodevelopmental Delay Enhances Kv12.2 Channel Inactivation.Int J Mol Sci. 2025 May 13;26(10):4631. doi: 10.3390/ijms26104631. Int J Mol Sci. 2025. PMID: 40429775 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
