

Neuronal actin cytoskeleton gain of function in the human brain
- PMID: 37517100
- PMCID: PMC10404607
- DOI: 10.1016/j.ebiom.2023.104725
Neuronal actin cytoskeleton gain of function in the human brain
Abstract
Background: While advancements in imaging techniques have led to major strides in deciphering the human brain, successful interventions are elusive and represent some of the most persistent translational gaps in medicine. Human restricted CHRFAM7A has been associated with neuropsychiatric disorders.
Methods: The physiological role of CHRFAM7A in human brain is explored using multiomics approach on 600 post mortem human brain tissue samples. The emerging pathways and mechanistic hypotheses are tested and validated in an isogenic hiPSC model of CHRFAM7A knock-in medial ganglionic eminence progenitors and neurons.
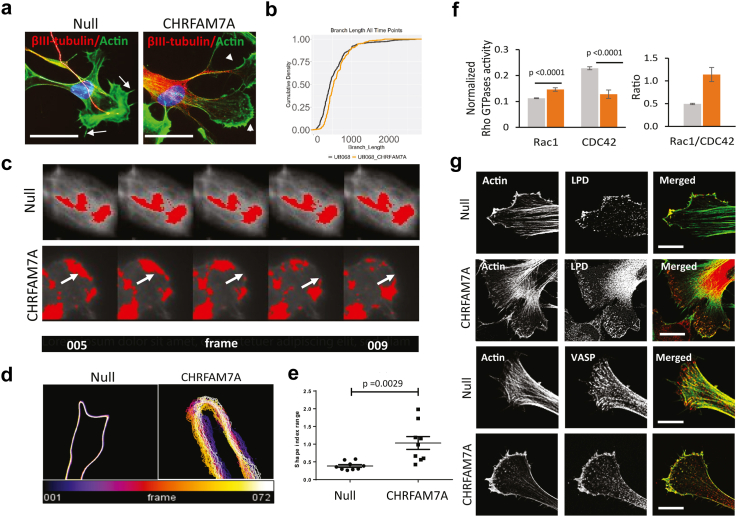
Findings: CHRFAM7A is identified as a modulator of intracellular calcium dynamics and an upstream regulator of Rac1. Rac1 activation re-designs the actin cytoskeleton leading to dynamic actin driven remodeling of membrane protrusion and a switch from filopodia to lamellipodia. The reinforced cytoskeleton leads to an advantage to tolerate stiffer mechanical properties of the extracellular environment.
Interpretation: CHRFAM7A modifies the actin cytoskeleton to a more dynamic and stiffness resistant state in an α7nAChR dependent manner. CHRFAM7A may facilitate neuronal adaptation to changes in the brain environment in physiological and pathological conditions contributing to risk or recovery. Understanding how CHRFAM7A affects human brain requires human studies in the areas of memory formation and erasure, cognitive reserve, and neuronal plasticity.
Funding: This work is supported in part by the Community Foundation for Greater Buffalo (Kinga Szigeti). Also, in part by the International Society for Neurochemistry (ISN) and The Company of Biologists (Nicolas Rosas). ROSMAP is supported by NIA grants P30AG10161, P30AG72975, R01AG15819, R01AG17917. U01AG46152, and U01AG61356.
Keywords: Actin cytoskeleton; CHRFAM7A; Human brain; Multiomics analysis; iPSC.
Copyright © 2023 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflict of interest.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
