

Conformational changes in the essential E. coli septal cell wall synthesis complex suggest an activation mechanism
- PMID: 37524712
- PMCID: PMC10390529
- DOI: 10.1038/s41467-023-39921-4
Conformational changes in the essential E. coli septal cell wall synthesis complex suggest an activation mechanism
Abstract
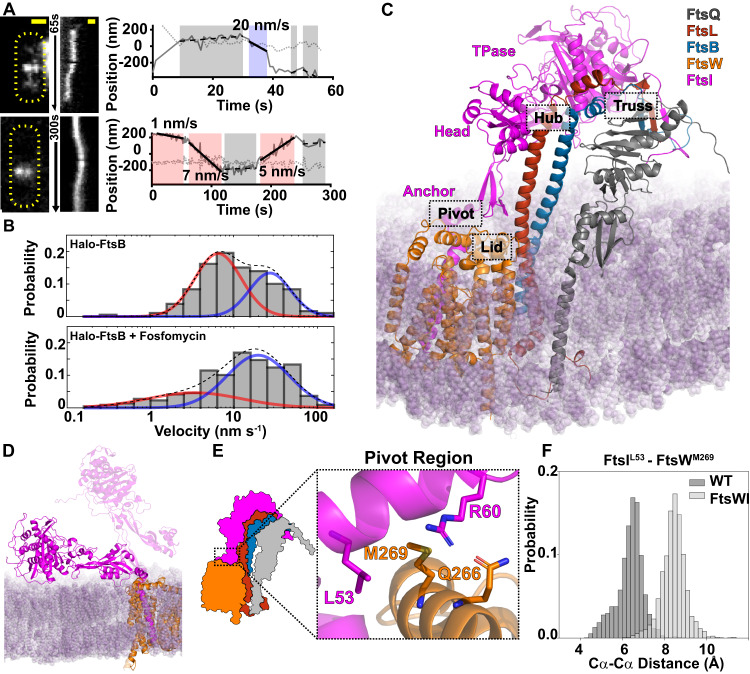
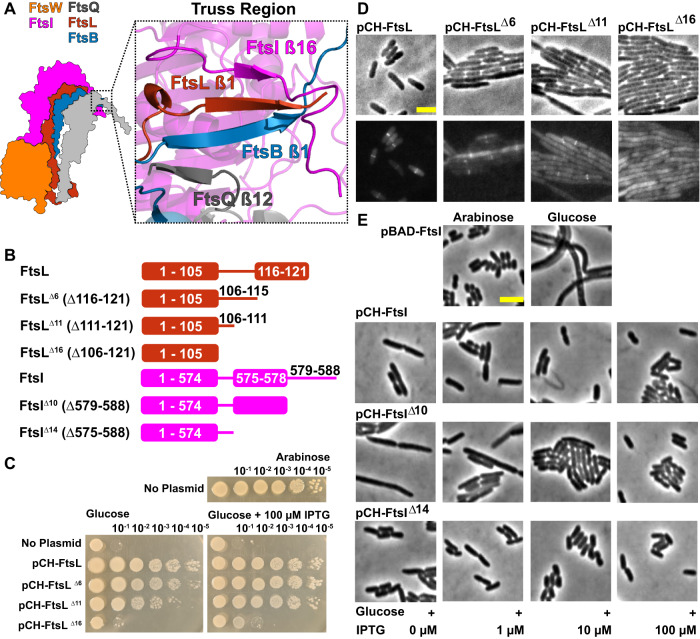
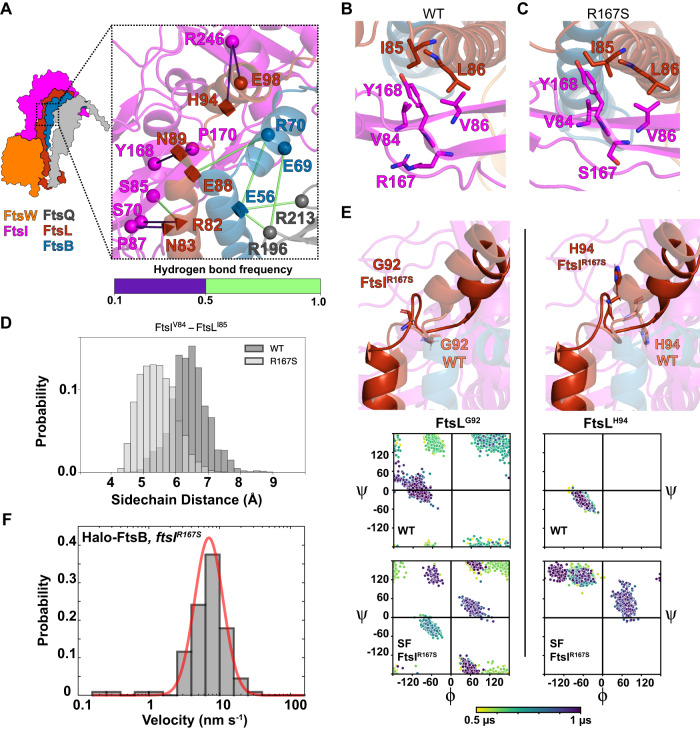
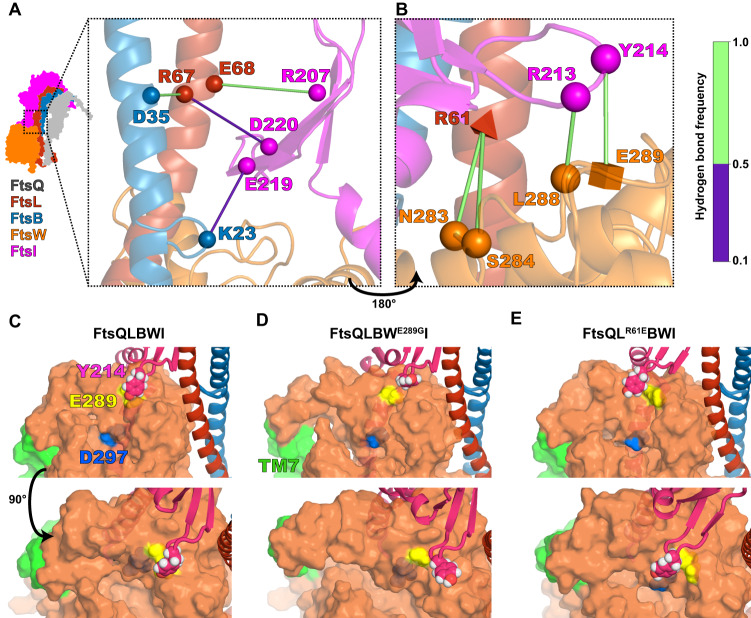
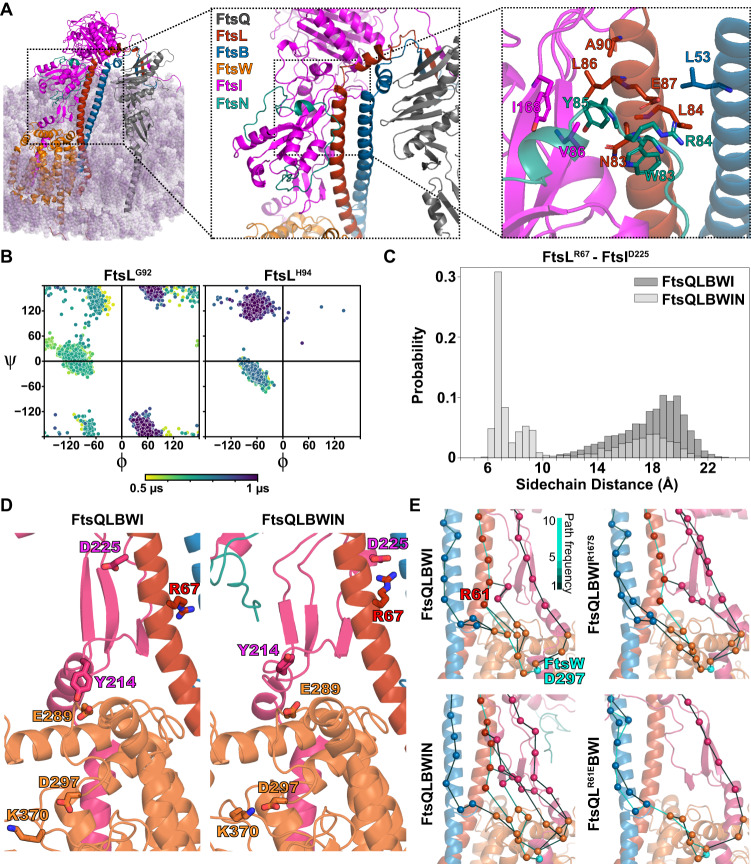
The bacterial divisome is a macromolecular machine composed of more than 30 proteins that controls cell wall constriction during division. Here, we present a model of the structure and dynamics of the core complex of the E. coli divisome, supported by a combination of structure prediction, molecular dynamics simulation, single-molecule imaging, and mutagenesis. We focus on the septal cell wall synthase complex formed by FtsW and FtsI, and its regulators FtsQ, FtsL, FtsB, and FtsN. The results indicate extensive interactions in four regions in the periplasmic domains of the complex. FtsQ, FtsL, and FtsB support FtsI in an extended conformation, with the FtsI transpeptidase domain lifted away from the membrane through interactions among the C-terminal domains. FtsN binds between FtsI and FtsL in a region rich in residues with superfission (activating) and dominant negative (inhibitory) mutations. Mutagenesis experiments and simulations suggest that the essential domain of FtsN links FtsI and FtsL together, potentially modulating interactions between the anchor-loop of FtsI and the putative catalytic cavity of FtsW, thus suggesting a mechanism of how FtsN activates the cell wall synthesis activities of FtsW and FtsI.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
FtsQ, FtsL and FtsI require FtsK, but not FtsN, for co-localization with FtsZ during Escherichia coli cell division.Mol Microbiol. 2001 Oct;42(2):395-413. doi: 10.1046/j.1365-2958.2001.02640.x. Mol Microbiol. 2001. PMID: 11703663
-
Structure of the heterotrimeric membrane protein complex FtsB-FtsL-FtsQ of the bacterial divisome.Nat Commun. 2023 Apr 5;14(1):1903. doi: 10.1038/s41467-023-37543-4. Nat Commun. 2023. PMID: 37019934 Free PMC article.
-
FtsA acts through FtsW to promote cell wall synthesis during cell division in Escherichia coli.Proc Natl Acad Sci U S A. 2021 Aug 31;118(35):e2107210118. doi: 10.1073/pnas.2107210118. Proc Natl Acad Sci U S A. 2021. PMID: 34453005 Free PMC article.
-
An Updated Model of the Divisome: Regulation of the Septal Peptidoglycan Synthesis Machinery by the Divisome.Int J Mol Sci. 2022 Mar 24;23(7):3537. doi: 10.3390/ijms23073537. Int J Mol Sci. 2022. PMID: 35408901 Free PMC article. Review.
-
Super-resolution images of peptidoglycan remodelling enzymes at the division site of Escherichia coli.Curr Genet. 2019 Feb;65(1):99-101. doi: 10.1007/s00294-018-0869-x. Epub 2018 Jul 28. Curr Genet. 2019. PMID: 30056491 Free PMC article. Review.
Cited by
-
Role of the antiparallel double-stranded filament form of FtsA in activating the Escherichia coli divisome.mBio. 2024 Aug 14;15(8):e0168724. doi: 10.1128/mbio.01687-24. Epub 2024 Jul 23. mBio. 2024. PMID: 39041810 Free PMC article.
-
The structural integrity of the membrane-embedded bacterial division complex FtsQBL studied with molecular dynamics simulations.Comput Struct Biotechnol J. 2023 Apr 3;21:2602-2612. doi: 10.1016/j.csbj.2023.03.052. eCollection 2023. Comput Struct Biotechnol J. 2023. PMID: 37114213 Free PMC article.
-
Regulating cellular metabolism and morphology to achieve high-yield synthesis of hyaluronan with controllable molecular weights.Nat Commun. 2025 Feb 28;16(1):2076. doi: 10.1038/s41467-025-56950-3. Nat Commun. 2025. PMID: 40021631 Free PMC article.
-
The divisome is a self-enhancing machine in Escherichia coli and Caulobacter crescentus.Nat Commun. 2024 Sep 18;15(1):8198. doi: 10.1038/s41467-024-52217-5. Nat Commun. 2024. PMID: 39294118 Free PMC article.
-
FtsZ-mediated spatial-temporal control over septal cell wall synthesis.Proc Natl Acad Sci U S A. 2025 Jul 8;122(27):e2426431122. doi: 10.1073/pnas.2426431122. Epub 2025 Jun 30. Proc Natl Acad Sci U S A. 2025. PMID: 40587787 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
