

Expanded olfactory system in ray-finned fishes capable of terrestrial exploration
- PMID: 37525196
- PMCID: PMC10392011
- DOI: 10.1186/s12915-023-01661-8
Expanded olfactory system in ray-finned fishes capable of terrestrial exploration
Abstract
Background: Smell abilities differ greatly among vertebrate species due to distinct sensory needs, with exceptional variability reported in the number of olfactory genes and the size of the odour-processing regions of the brain. However, key environmental factors shaping genomic and phenotypic changes linked to the olfactory system remain difficult to identify at macroevolutionary scales. Here, we investigate the association between diverse ecological traits and the number of olfactory chemoreceptors in approximately two hundred ray-finned fishes.
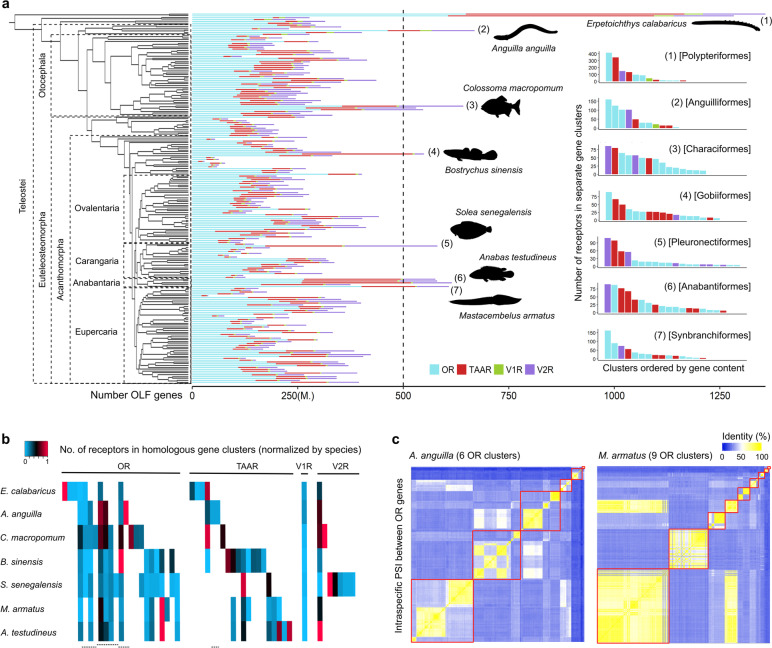
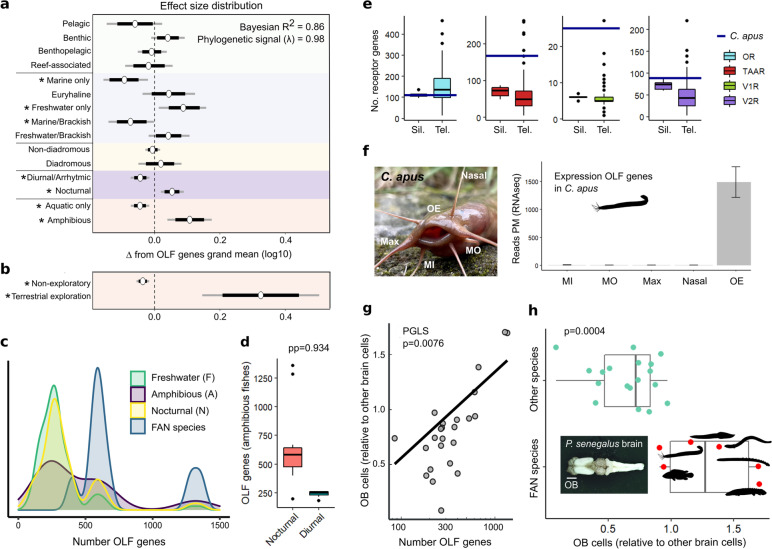
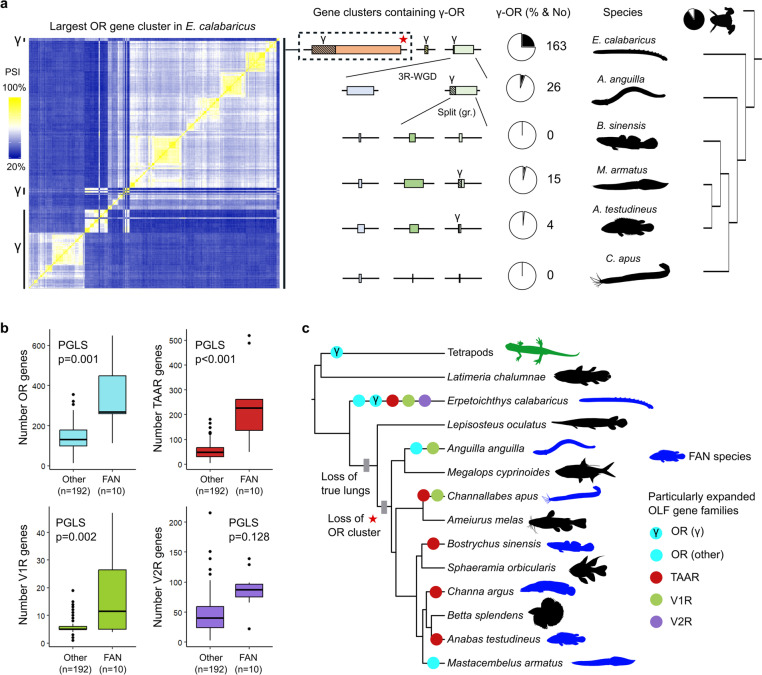
Results: We found independent expansions producing large gene repertoires in several lineages of nocturnal amphibious fishes, generally able to perform active terrestrial exploration. We reinforced this finding with on-purpose genomic and transcriptomic analysis of Channallabes apus, a catfish species from a clade with chemosensory-based aerial orientation. Furthermore, we also detected an augmented information-processing capacity in the olfactory bulb of nocturnal amphibious fishes by estimating the number of cells contained in this brain region in twenty-four actinopterygian species.
Conclusions: Overall, we report a convergent genomic and phenotypic magnification of the olfactory system in nocturnal amphibious fishes. This finding suggests the possibility of an analogous evolutionary event in fish-like tetrapod ancestors during the first steps of the water-to-land transition, favouring terrestrial adaptation through enhanced aerial orientation.
Keywords: Amphibious fishes; Evolutionary transition; Olfactory receptors; Sensory evolution.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Iwaniuk AN. 1.18 - Functional Correlates of Brain and Brain Region Sizes in Nonmammalian Vertebrates. In: Kaas JHBT-E of NS, editor. 2nd ed. Oxford: Academic Press; 2017. p. 335–48.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
