

The induction of p53 correlates with defects in the production, but not the levels, of the small ribosomal subunit and stalled large ribosomal subunit biogenesis
- PMID: 37526268
- PMCID: PMC10516649
- DOI: 10.1093/nar/gkad637
The induction of p53 correlates with defects in the production, but not the levels, of the small ribosomal subunit and stalled large ribosomal subunit biogenesis
Abstract
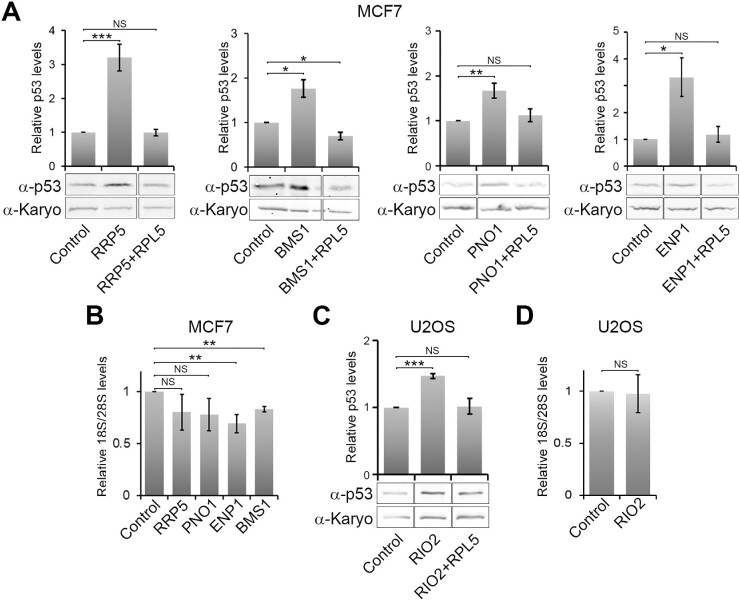
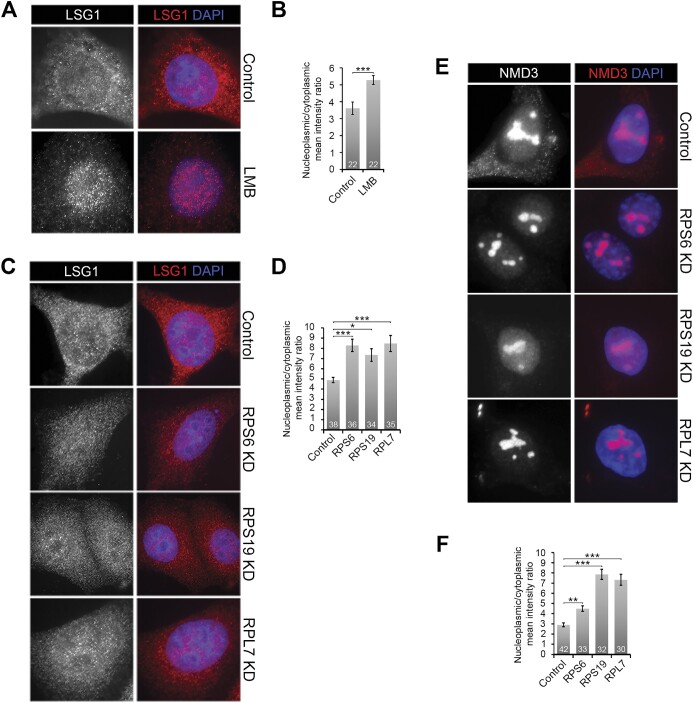
Ribosome biogenesis is one of the biggest consumers of cellular energy. More than 20 genetic diseases (ribosomopathies) and multiple cancers arise from defects in the production of the 40S (SSU) and 60S (LSU) ribosomal subunits. Defects in the production of either the SSU or LSU result in p53 induction through the accumulation of the 5S RNP, an LSU assembly intermediate. While the mechanism is understood for the LSU, it is still unclear how SSU production defects induce p53 through the 5S RNP since the production of the two subunits is believed to be uncoupled. Here, we examined the response to SSU production defects to understand how this leads to the activation of p53 via the 5S RNP. We found that p53 activation occurs rapidly after SSU production is blocked, prior to changes in mature ribosomal RNA (rRNA) levels but correlated with early, middle and late SSU pre-rRNA processing defects. Furthermore, both nucleolar/nuclear LSU maturation, in particular late stages in 5.8S rRNA processing, and pre-LSU export were affected by SSU production defects. We have therefore uncovered a novel connection between the SSU and LSU production pathways in human cells, which explains how p53 is induced in response to SSU production defects.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures






Similar articles
-
The nucleolar protein NOL12 is required for processing of large ribosomal subunit rRNA precursors in Arabidopsis.BMC Plant Biol. 2023 Nov 3;23(1):538. doi: 10.1186/s12870-023-04561-9. BMC Plant Biol. 2023. PMID: 37919659 Free PMC article.
-
The roles of RRP15 in nucleolar formation, ribosome biogenesis and checkpoint control in human cells.Oncotarget. 2017 Feb 21;8(8):13240-13252. doi: 10.18632/oncotarget.14658. Oncotarget. 2017. PMID: 28099941 Free PMC article.
-
Assembly and structure of the SSU processome-a nucleolar precursor of the small ribosomal subunit.Curr Opin Struct Biol. 2018 Apr;49:85-93. doi: 10.1016/j.sbi.2018.01.008. Epub 2018 Feb 4. Curr Opin Struct Biol. 2018. PMID: 29414516 Free PMC article. Review.
-
The RNA recognition motif of NIFK is required for rRNA maturation during cell cycle progression.RNA Biol. 2015;12(3):255-67. doi: 10.1080/15476286.2015.1017221. RNA Biol. 2015. PMID: 25826659 Free PMC article.
-
Ribosomal Proteins Control Tumor Suppressor Pathways in Response to Nucleolar Stress.Bioessays. 2019 Mar;41(3):e1800183. doi: 10.1002/bies.201800183. Epub 2019 Feb 1. Bioessays. 2019. PMID: 30706966 Review.
Cited by
-
Ribosome-directed cancer therapies: the tip of the iceberg?Trends Pharmacol Sci. 2025 Apr;46(4):303-310. doi: 10.1016/j.tips.2025.02.001. Epub 2025 Mar 4. Trends Pharmacol Sci. 2025. PMID: 40044536 Review.
-
p53 in the Molecular Circuitry of Bone Marrow Failure Syndromes.Int J Mol Sci. 2023 Oct 6;24(19):14940. doi: 10.3390/ijms241914940. Int J Mol Sci. 2023. PMID: 37834388 Free PMC article. Review.
-
The Effects of Deregulated Ribosomal Biogenesis in Cancer.Biomolecules. 2023 Oct 30;13(11):1593. doi: 10.3390/biom13111593. Biomolecules. 2023. PMID: 38002277 Free PMC article. Review.
-
Potentials of ribosomopathy gene as pharmaceutical targets for cancer treatment.J Pharm Anal. 2024 Mar;14(3):308-320. doi: 10.1016/j.jpha.2023.10.001. Epub 2023 Oct 13. J Pharm Anal. 2024. PMID: 38618250 Free PMC article. Review.
-
The Beak of Eukaryotic Ribosomes: Life, Work and Miracles.Biomolecules. 2024 Jul 22;14(7):882. doi: 10.3390/biom14070882. Biomolecules. 2024. PMID: 39062596 Free PMC article. Review.
References
-
- Lempiainen H., Shore D. Growth control and ribosome biogenesis. Curr. Opin. Cell Biol. 2009; 21:855–863. - PubMed
-
- Teng T., Thomas G., Mercer C.A.. Growth control and ribosomopathies. Curr. Opin. Genet. Dev. 2013; 23:63–71. - PubMed
-
- Warner J.R. The economics of ribosome biosynthesis in yeast. Trends Biochem. Sci. 1999; 24:437–440. - PubMed
-
- Bustelo X.R., Dosil M.. Ribosome biogenesis and cancer: basic and translational challenges. Curr. Opin. Genet. Dev. 2018; 48:22–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
