

Adipose METTL14-Elicited N6 -Methyladenosine Promotes Obesity, Insulin Resistance, and NAFLD Through Suppressing β Adrenergic Signaling and Lipolysis
- PMID: 37526326
- PMCID: PMC10558699
- DOI: 10.1002/advs.202301645
Adipose METTL14-Elicited N6 -Methyladenosine Promotes Obesity, Insulin Resistance, and NAFLD Through Suppressing β Adrenergic Signaling and Lipolysis
Abstract
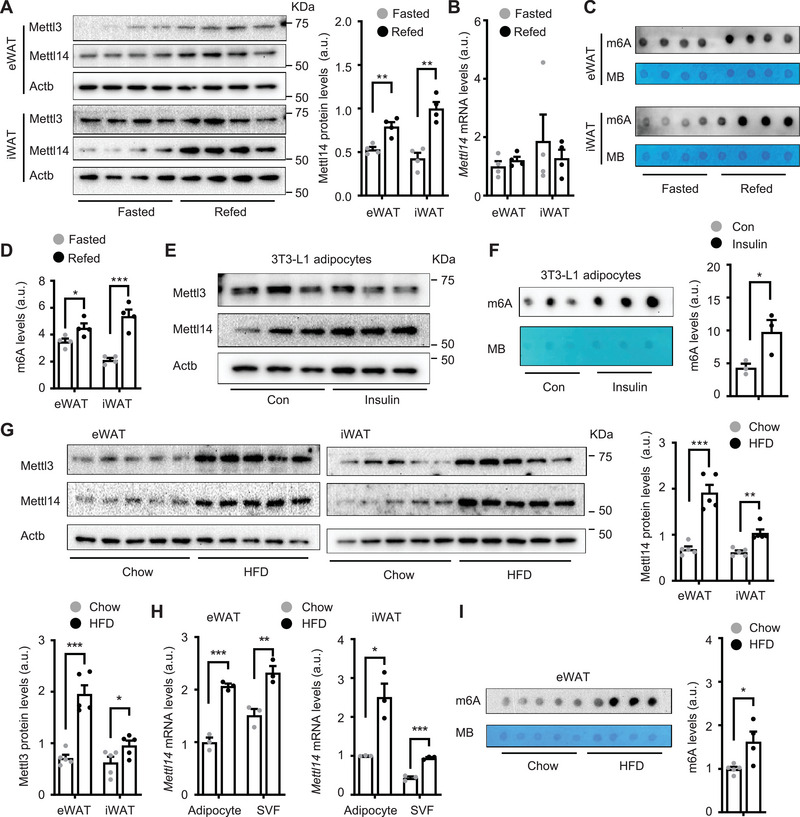
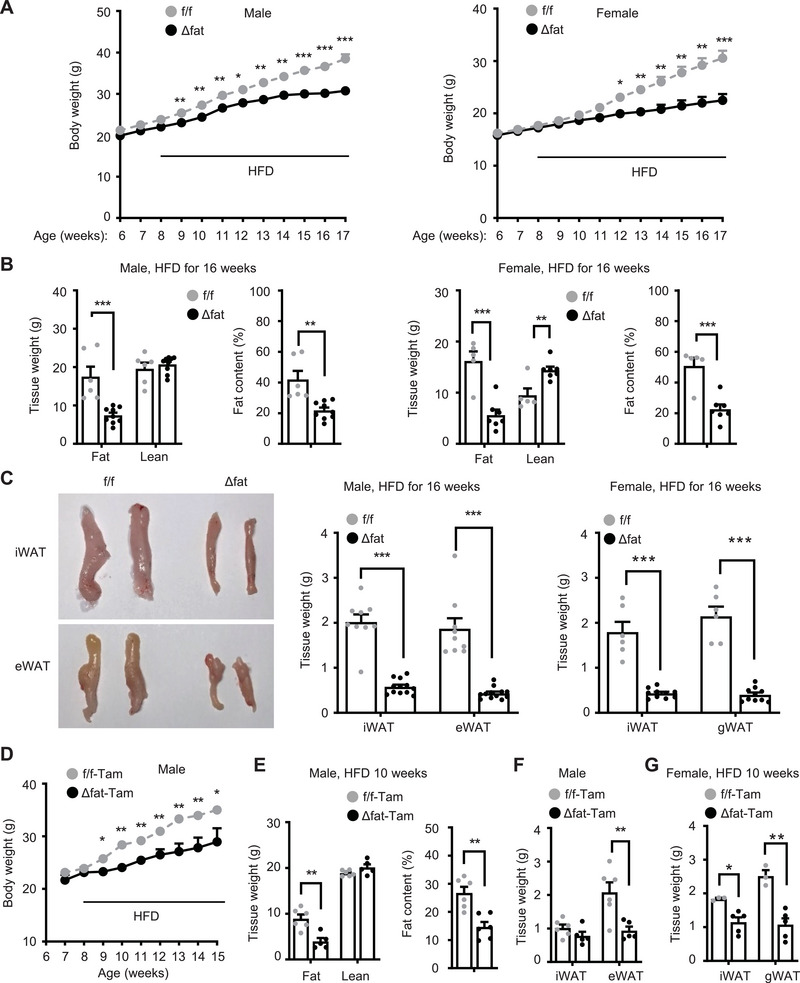
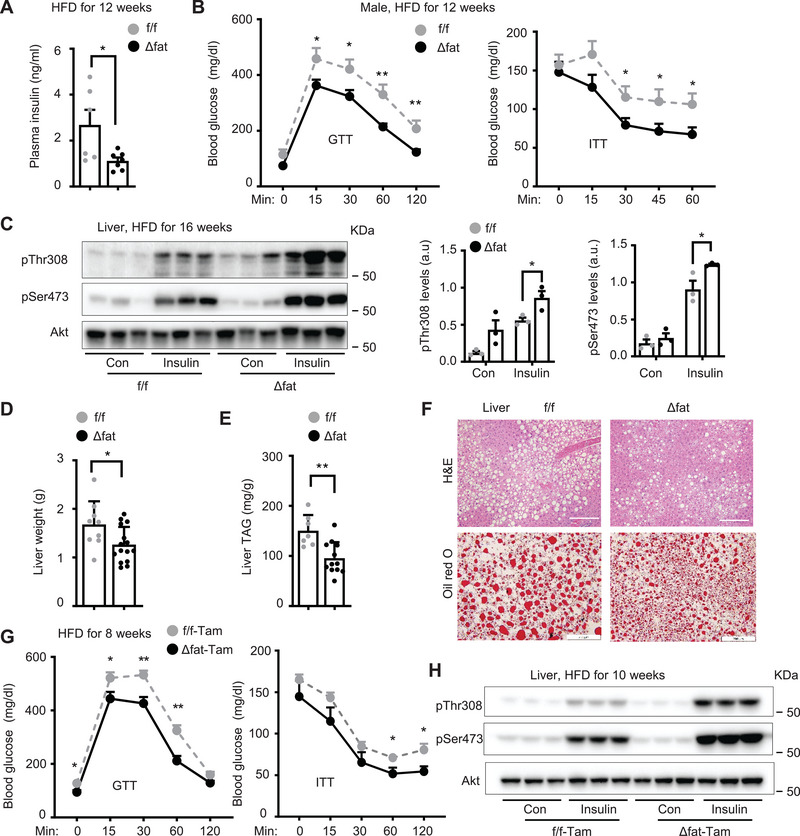
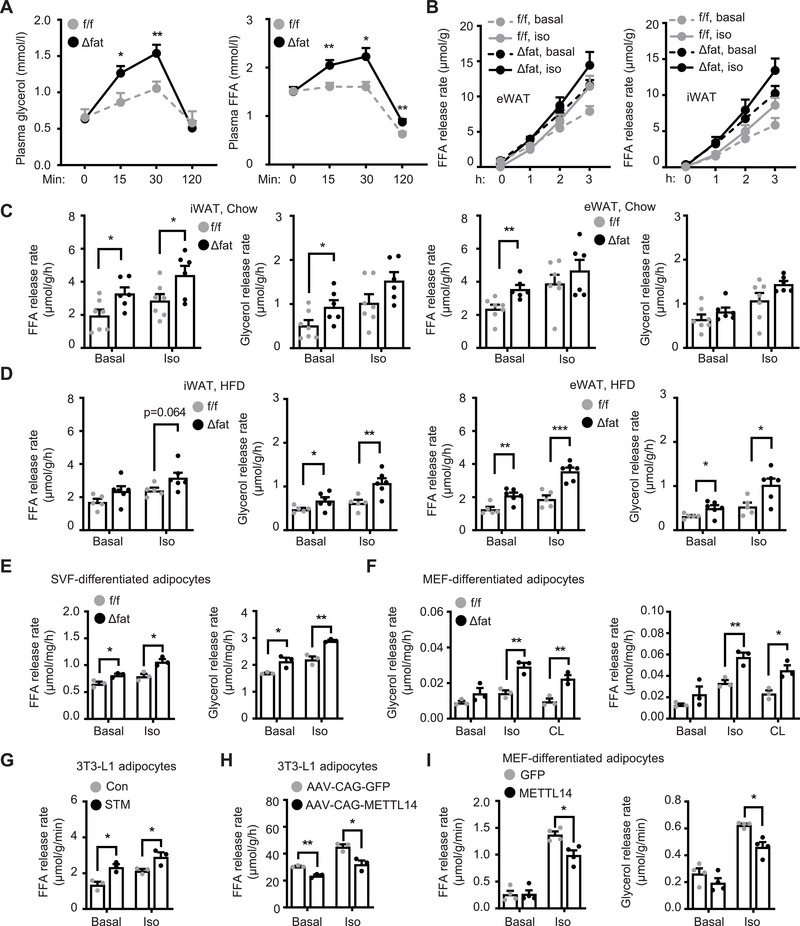
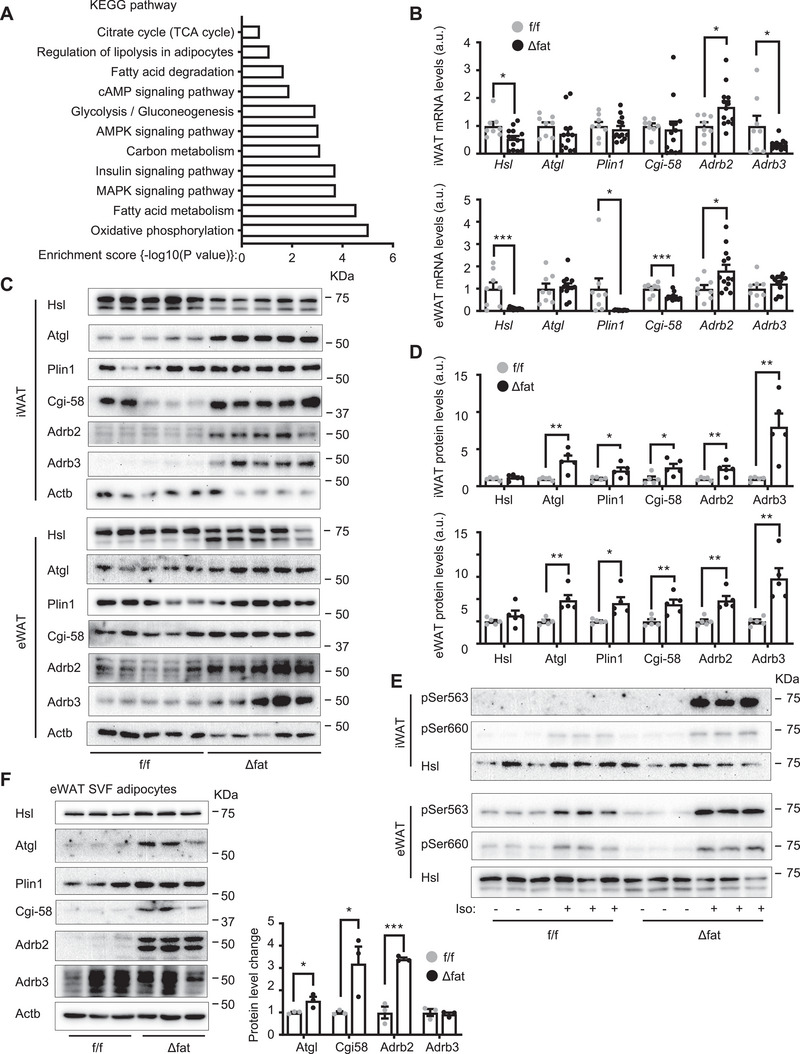
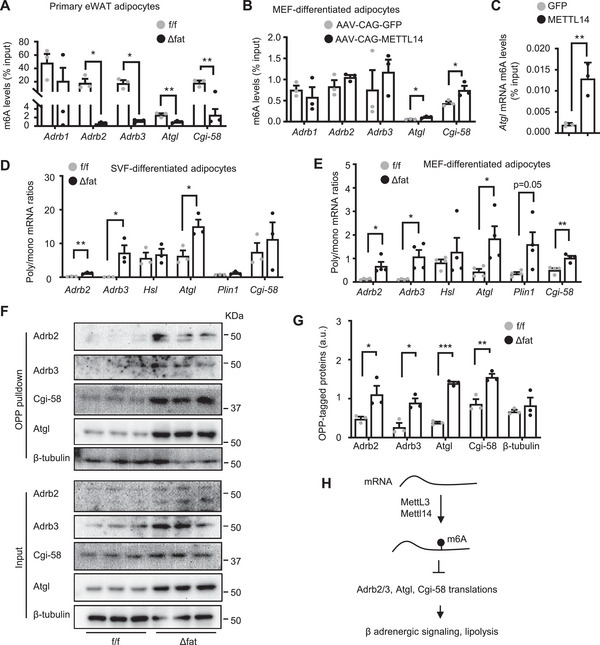
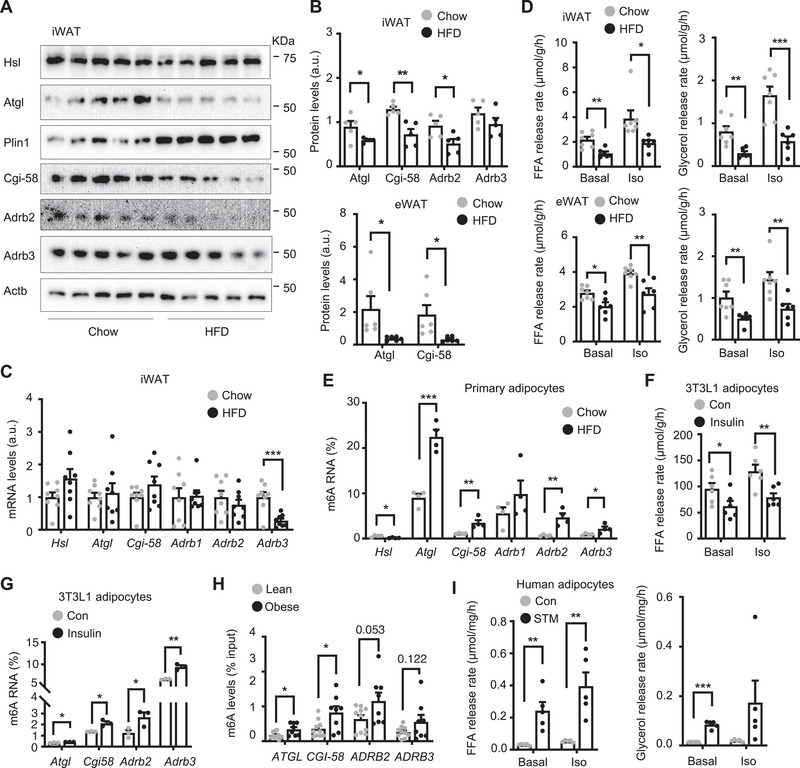
White adipose tissue (WAT) lipolysis releases free fatty acids as a key energy substance to support metabolism in fasting, cold exposure, and exercise. Atgl, in concert with Cgi-58, catalyzes the first lipolytic reaction. The sympathetic nervous system (SNS) stimulates lipolysis via neurotransmitter norepinephrine that activates adipocyte β adrenergic receptors (Adrb1-3). In obesity, adipose Adrb signaling and lipolysis are impaired, contributing to pathogenic WAT expansion; however, the underling mechanism remains poorly understood. Recent studies highlight importance of N6 -methyladenosine (m6A)-based RNA modification in health and disease. METTL14 heterodimerizes with METTL3 to form an RNA methyltransferase complex that installs m6A in transcripts. Here, this work shows that adipose Mettl3 and Mettl14 are influenced by fasting, refeeding, and insulin, and are upregulated in high fat diet (HFD) induced obesity. Adipose Adrb2, Adrb3, Atgl, and Cgi-58 transcript m6A contents are elevated in obesity. Mettl14 ablation decreases these transcripts' m6A contents and increases their translations and protein levels in adipocytes, thereby increasing Adrb signaling and lipolysis. Mice with adipocyte-specific deletion of Mettl14 are resistant to HFD-induced obesity, insulin resistance, glucose intolerance, and nonalcoholic fatty liver disease (NAFLD). These results unravel a METTL14/m6A/translation pathway governing Adrb signaling and lipolysis. METTL14/m6A-based epitranscriptomic reprogramming impairs adipose Adrb signaling and lipolysis, promoting obesity, NAFLD, and metabolic disease.
Keywords: RNA modifications; adipose tissue; diabetes; lipolysis; m6A; obesity; β-adrenergic signaling.
© 2023 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- a) Sun C., Jiang L., Liu Y., Shen H., Weiss S. J., Zhou Y., Rui L., Cell Rep. 2016, 17, 2015; - PMC - PubMed
- b) Ahmadian M., Abbott M. J., Tang T., Hudak C. S., Kim Y., Bruss M., Hellerstein M. K., Lee H. Y., Samuel V. T., Shulman G. I., Wang Y., Duncan R. E., Kang C., Sul H. S., Cell Metab. 2011, 13, 739; - PMC - PubMed
- c) Wu J. W., Wang S. P., Casavant S., Moreau A., Yang G. S., Mitchell G. A., Endocrinology 2012, 153, 2198. - PubMed
-
- a) Ahmadian M., Duncan R. E., Varady K. A., Frasson D., Hellerstein M. K., Birkenfeld A. L., Samuel V. T., Shulman G. I., Wang Y., Kang C., Sul H. S., Diabetes 2009, 58, 855; - PMC - PubMed
- b) Zhang X., Xie X., Heckmann B. L., Saarinen A. M., Czyzyk T. A., Liu J., Diabetes 2014, 63, 934; - PMC - PubMed
- c) El‐Assaad W., El‐Kouhen K., Mohammad A. H., Yang J., Morita M., Gamache I., Mamer O., Avizonis D., Hermance N., Kersten S., Tremblay M. L., Kelliher M. A., Teodoro J. G., Diabetologia 2015, 58, 149. - PMC - PubMed
-
- a) Pirzgalska R. M., Seixas E., Seidman J. S., Link V. M., Sanchez N. M., Mahu I., Mendes R., Gres V., Kubasova N., Morris I., Arus B. A., Larabee C. M., Vasques M., Tortosa F., Sousa A. L., Anandan S., Tranfield E., Hahn M. K., Iannacone M., Spann N. J., Glass C. K., Domingos A. I., Nat. Med. 2017, 23, 1309; - PMC - PubMed
- b) Camell C. D., Sander J., Spadaro O., Lee A., Nguyen K. Y., Wing A., Goldberg E. L., Youm Y. H., Brown C. W., Elsworth J., Rodeheffer M. S., Schultze J. L., Dixit V. D., Nature 2017, 550, 119; - PMC - PubMed
- c) Gao H., Arner P., Beauchef G., Guere C., Vie K., Dahlman I., Mejhert N., Ryden M., Cell Metab. 2020, 32, 1; - PubMed
- d) Arner P., Bernard S., Appelsved L., Fu K. Y., Andersson D. P., Salehpour M., Thorell A., Ryden M., Spalding K. L., Nat. Med. 2019, 25, 1385; - PubMed
- e) Villena J. A., Roy S., Sarkadi‐Nagy E., Kim K. H., Sul H. S., J. Biol. Chem. 2004, 279, 47066; - PubMed
- f) Kim J. Y., Tillison K., Lee J. H., Rearick D. A., Smas C. M., Am. J. Physiol. Endocrinol. Metab. 2006, 291, E115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK125513/DK/NIDDK NIH HHS/United States
- R01 DK130111/DK/NIDDK NIH HHS/United States
- R56 DK125513/DK/NIDDK NIH HHS/United States
- R01 DK130875/DK/NIDDK NIH HHS/United States
- R01 DK127568/DK/NIDDK NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- R56 DK132785/DK/NIDDK NIH HHS/United States
- R01 DK121759/DK/NIDDK NIH HHS/United States
- P30 DK089503/DK/NIDDK NIH HHS/United States
- I01 CX001811/CX/CSRD VA/United States
- P60 DK020572/DK/NIDDK NIH HHS/United States
- P30 DK034933/DK/NIDDK NIH HHS/United States
- R01 DK114220/DK/NIDDK NIH HHS/United States
- NH/NIH HHS/United States
- P30 DK020595/DK/NIDDK NIH HHS/United States
- U2C DK110768/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical