

Central role of Tim17 in mitochondrial presequence protein translocation
- PMID: 37527780
- PMCID: PMC10511324
- DOI: 10.1038/s41586-023-06477-8
Central role of Tim17 in mitochondrial presequence protein translocation
Abstract
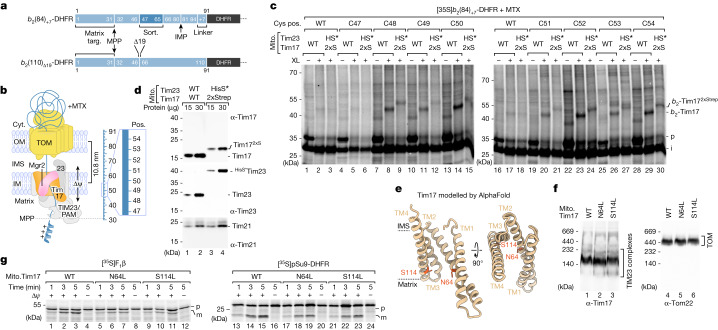
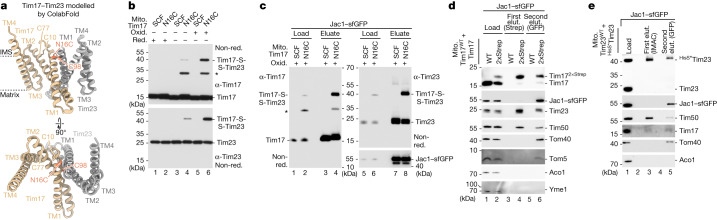
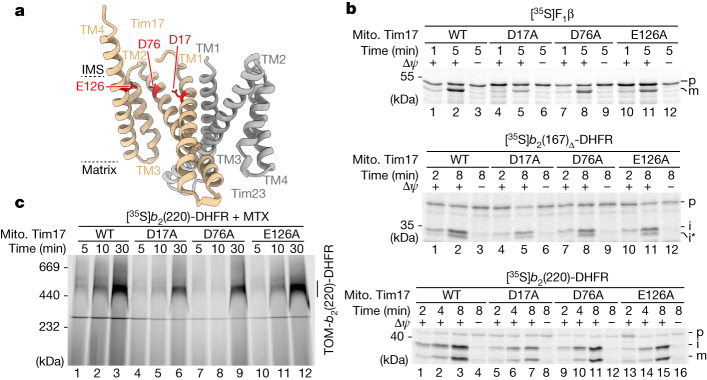
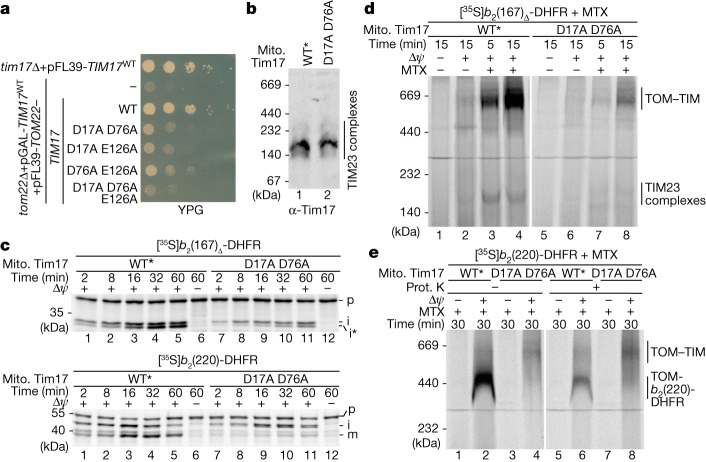
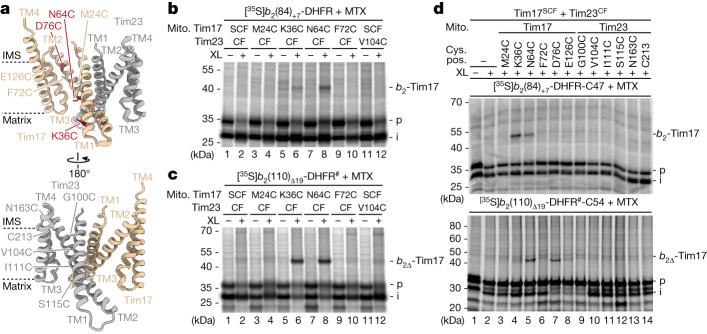
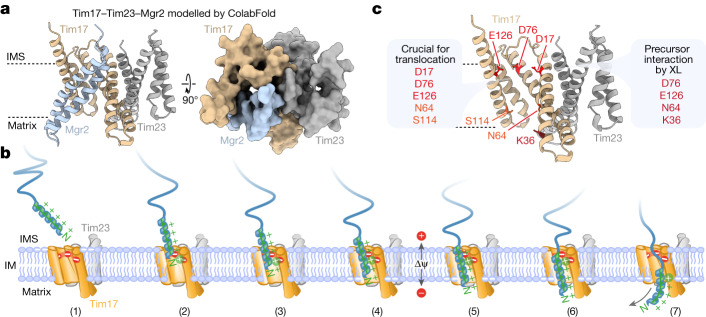
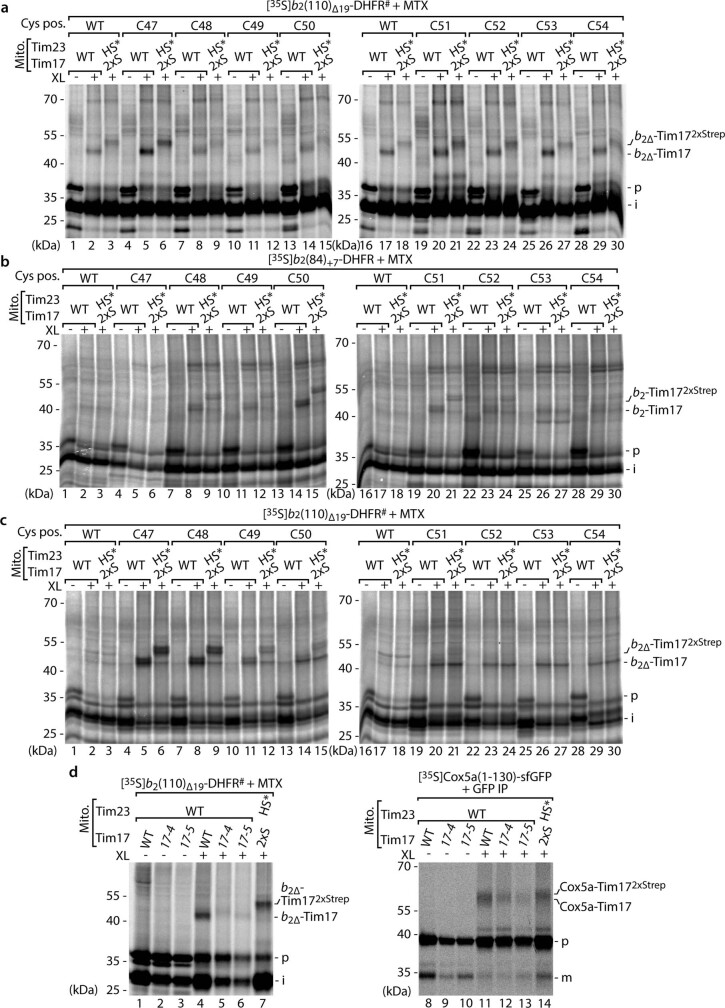
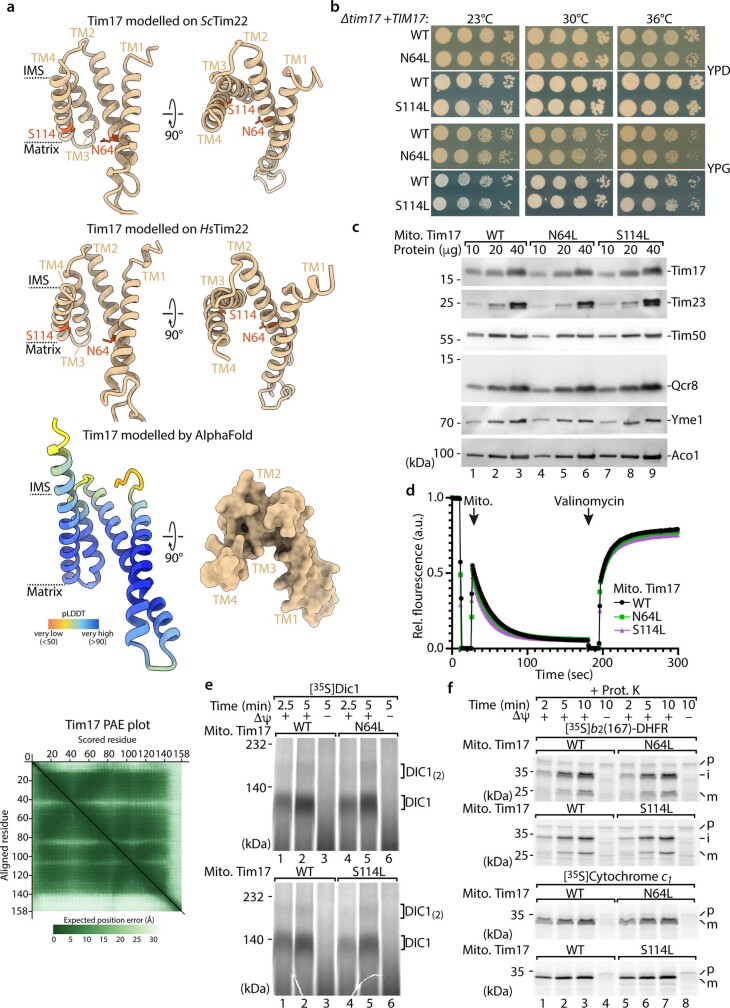
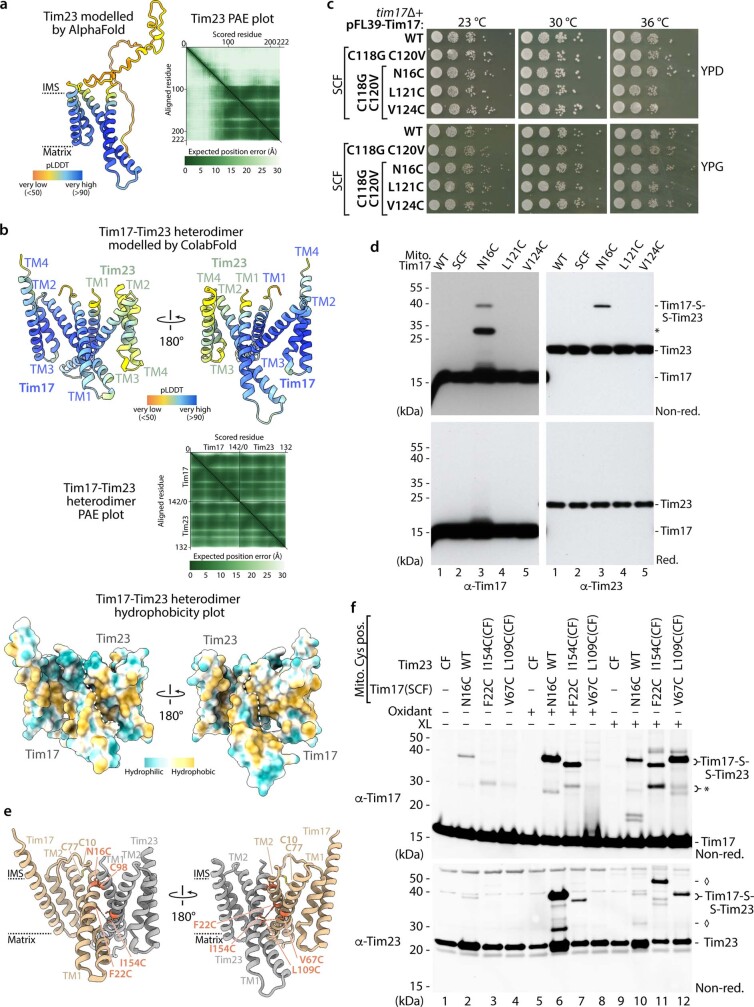
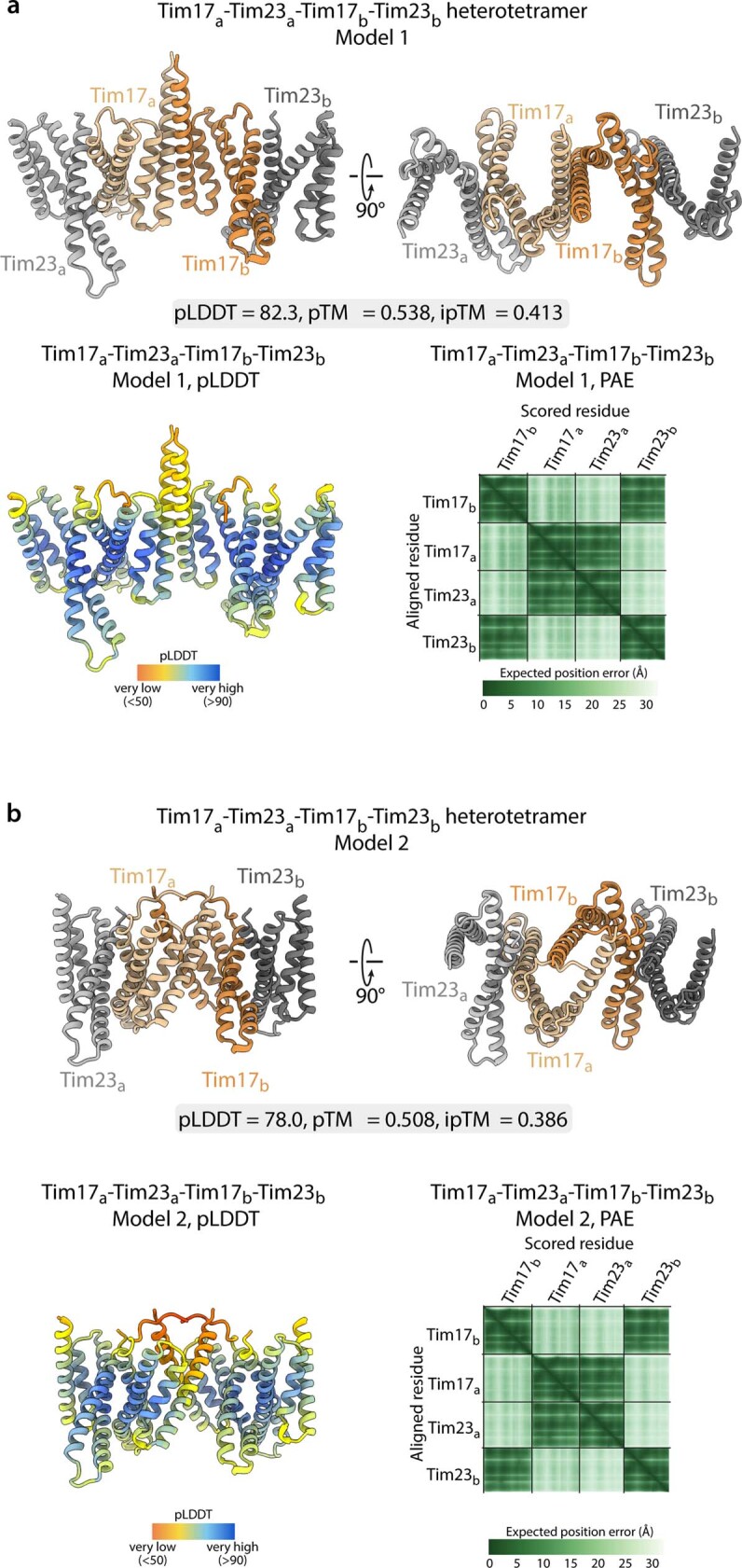
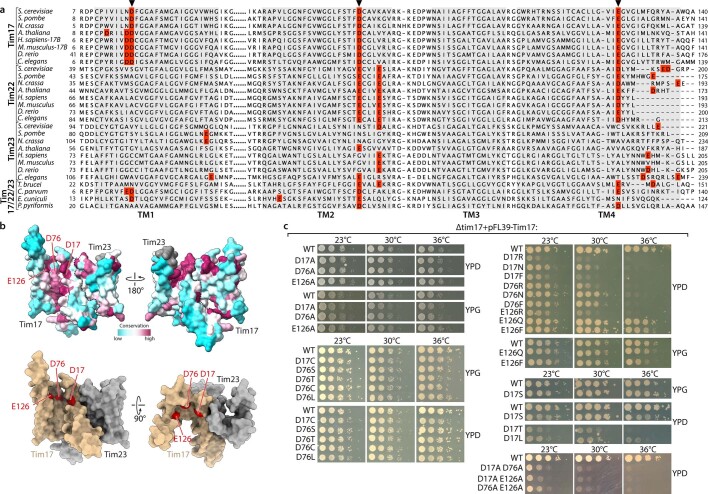
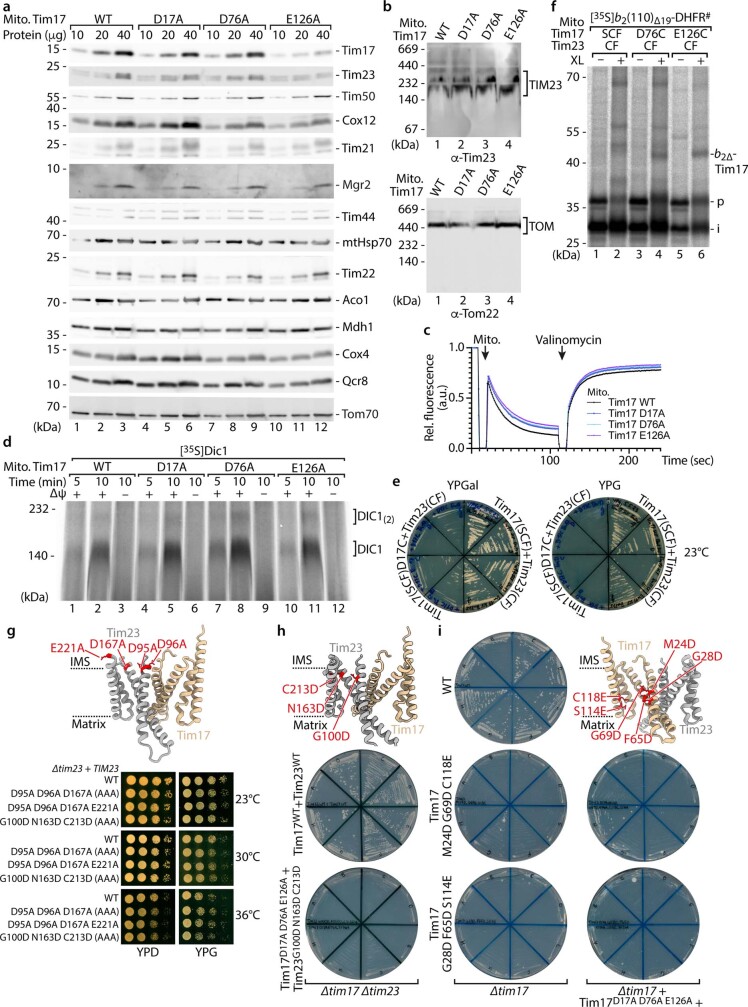
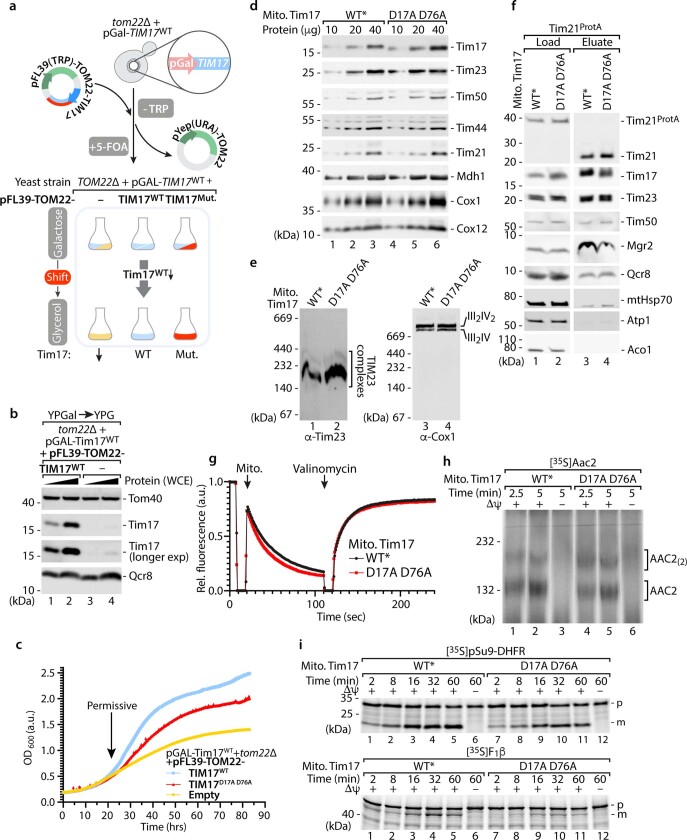
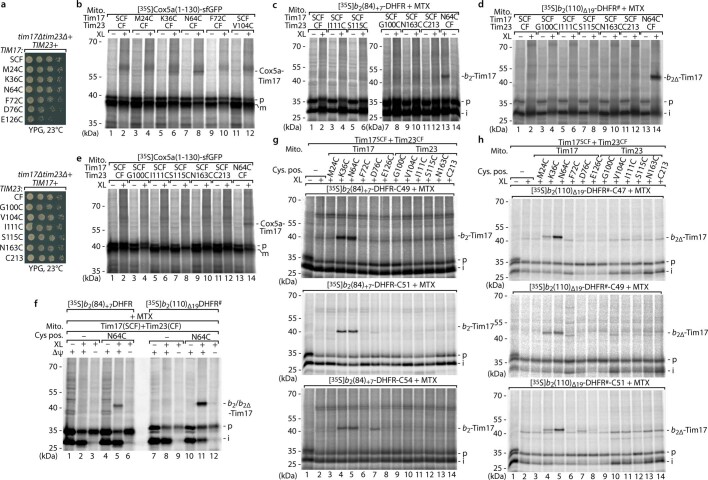
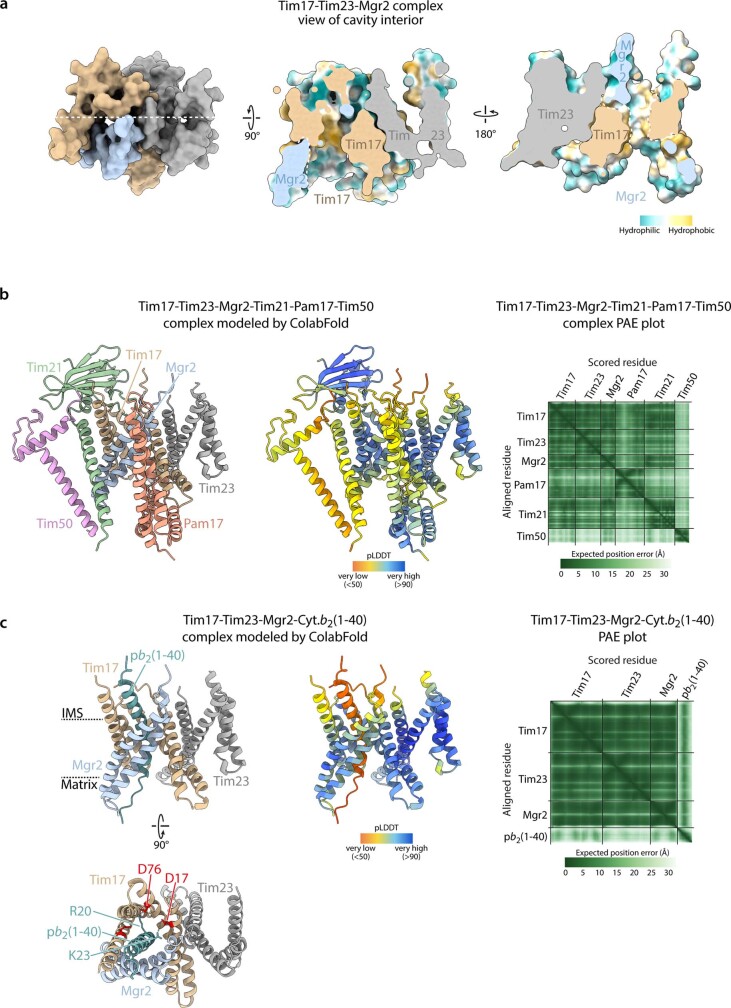
The presequence translocase of the mitochondrial inner membrane (TIM23) represents the major route for the import of nuclear-encoded proteins into mitochondria1,2. About 60% of more than 1,000 different mitochondrial proteins are synthesized with amino-terminal targeting signals, termed presequences, which form positively charged amphiphilic α-helices3,4. TIM23 sorts the presequence proteins into the inner membrane or matrix. Various views, including regulatory and coupling functions, have been reported on the essential TIM23 subunit Tim17 (refs. 5-7). Here we mapped the interaction of Tim17 with matrix-targeted and inner membrane-sorted preproteins during translocation in the native membrane environment. We show that Tim17 contains conserved negative charges close to the intermembrane space side of the bilayer, which are essential to initiate presequence protein translocation along a distinct transmembrane cavity of Tim17 for both classes of preproteins. The amphiphilic character of mitochondrial presequences directly matches this Tim17-dependent translocation mechanism. This mechanism permits direct lateral release of transmembrane segments of inner membrane-sorted precursors into the inner membrane.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
