

Peroxisome proliferator activated receptor-γ in osteoblasts controls bone formation and fat mass by regulating sclerostin expression
- PMID: 37534168
- PMCID: PMC10391670
- DOI: 10.1016/j.isci.2023.106999
Peroxisome proliferator activated receptor-γ in osteoblasts controls bone formation and fat mass by regulating sclerostin expression
Abstract
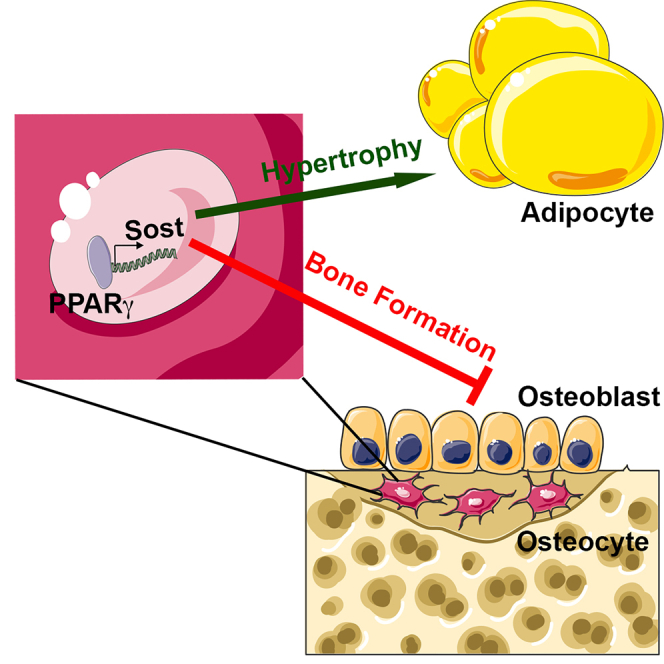
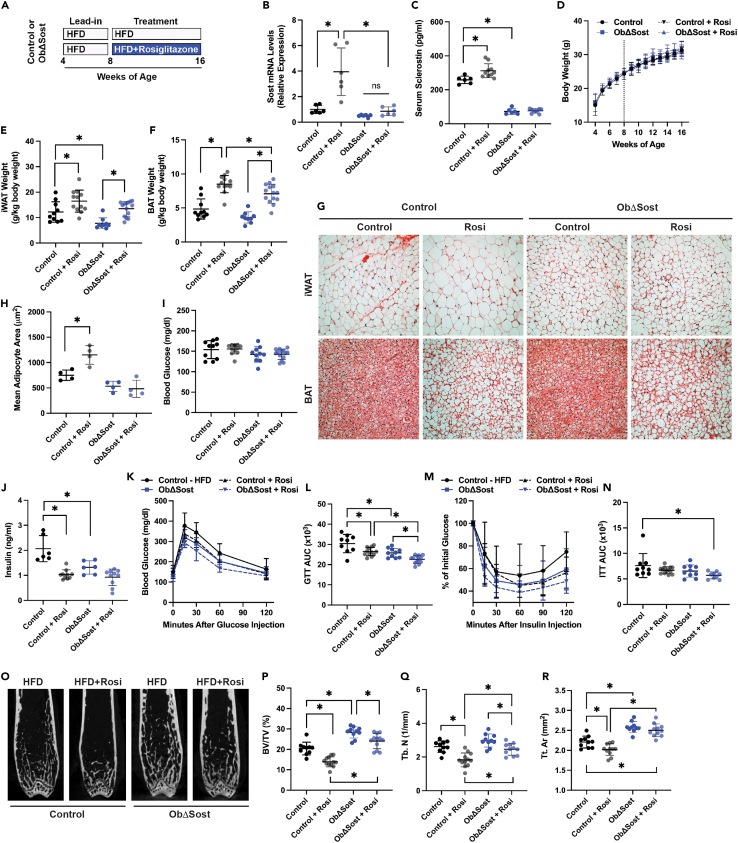
The nuclear receptor peroxisome proliferator activated receptor-γ (PPARγ) is a key contributor to metabolic function via its adipogenic and insulin-sensitizing functions, but it has negative effects on skeletal homeostasis. Here, we questioned whether the skeletal and metabolic actions of PPARγ are linked. Ablating Pparg expression in osteoblasts and osteocytes produced a high bone mass phenotype, secondary to increased osteoblast activity, and a reduction in subcutaneous fat mass because of reduced fatty acid synthesis and increased fat oxidation. The skeletal and metabolic phenotypes in Pparg mutants proceed from the regulation of sclerostin production by PPARγ. Mutants exhibited reductions in skeletal Sost expression and serum sclerostin levels while increasing production normalized both phenotypes. Importantly, disrupting the production of sclerostin synergized with the insulin-sensitizing actions of a PPARγ agonist while preventing bone loss. These data suggest that modulating sclerostin action may prevent bone loss associated with anti-diabetic therapies and augment their metabolic actions.
Keywords: Cell biology; Molecular physiology.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Figures




Similar articles
-
PPARG in osteocytes controls sclerostin expression, bone mass, marrow adiposity and mediates TZD-induced bone loss.Bone. 2021 Jun;147:115913. doi: 10.1016/j.bone.2021.115913. Epub 2021 Mar 16. Bone. 2021. PMID: 33722775 Free PMC article.
-
Conditional Deletion of Sost in MSC-Derived Lineages Identifies Specific Cell-Type Contributions to Bone Mass and B-Cell Development.J Bone Miner Res. 2018 Oct;33(10):1748-1759. doi: 10.1002/jbmr.3467. Epub 2018 Jun 29. J Bone Miner Res. 2018. PMID: 29750826
-
Partial agonist, telmisartan, maintains PPARγ serine 112 phosphorylation, and does not affect osteoblast differentiation and bone mass.PLoS One. 2014 May 8;9(5):e96323. doi: 10.1371/journal.pone.0096323. eCollection 2014. PLoS One. 2014. PMID: 24810249 Free PMC article.
-
Peroxisome proliferator-activated receptor gamma expression along the male genital system and its role in male fertility.Hum Reprod. 2020 Sep 1;35(9):2072-2085. doi: 10.1093/humrep/deaa153. Hum Reprod. 2020. PMID: 32766764 Review.
-
Role and mechanism of action of sclerostin in bone.Bone. 2017 Mar;96:29-37. doi: 10.1016/j.bone.2016.10.007. Epub 2016 Oct 12. Bone. 2017. PMID: 27742498 Free PMC article. Review.
Cited by
-
The role of bone in whole-body energy metabolism.Nat Rev Endocrinol. 2025 Aug 22. doi: 10.1038/s41574-025-01162-4. Online ahead of print. Nat Rev Endocrinol. 2025. PMID: 40846895 Review.
-
Regulatory Effect of Osteocytes on Extramedullary and Bone Marrow Adipose Tissue Development and Function.Curr Osteoporos Rep. 2024 Jun;22(3):301-307. doi: 10.1007/s11914-024-00871-5. Epub 2024 Apr 16. Curr Osteoporos Rep. 2024. PMID: 38625510 Free PMC article. Review.
-
PTH receptor signalling, osteocytes and bone disease induced by diabetes mellitus.Nat Rev Endocrinol. 2024 Nov;20(11):661-672. doi: 10.1038/s41574-024-01014-7. Epub 2024 Jul 17. Nat Rev Endocrinol. 2024. PMID: 39020007 Review.
References
-
- Kajimura D., Lee H.W., Riley K.J., Arteaga-Solis E., Ferron M., Zhou B., Clarke C.J., Hannun Y.A., DePinho R.A., Guo X.E., et al. Adiponectin regulates bone mass via opposite central and peripheral mechanisms through FoxO1. Cell Metab. 2013;17:901–915. doi: 10.1016/j.cmet.2013.04.009. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
