

Migratory songbirds exhibit seasonal modulation of the oxygen cascade
- PMID: 37534524
- PMCID: PMC10482389
- DOI: 10.1242/jeb.245975
Migratory songbirds exhibit seasonal modulation of the oxygen cascade
Abstract
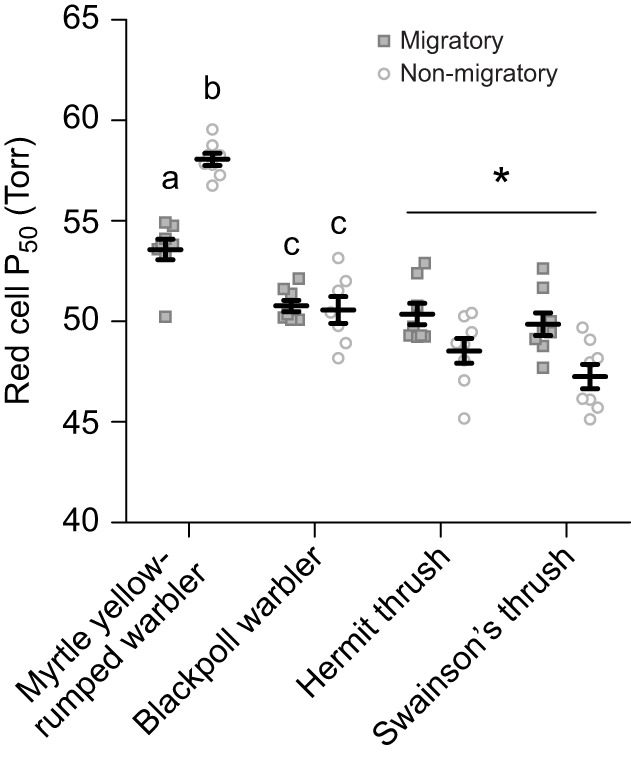
Migratory flight requires birds to maintain intensive aerobic exercise for many hours or days. Maintaining O2 supply to flight muscles is therefore important during migration, especially since migratory songbirds have been documented flying at altitudes greater than 5000 m above sea level, where O2 is limited. Whether songbirds exhibit seasonal plasticity of the O2 cascade to maintain O2 uptake and transport during migratory flight is not well understood. We investigated changes in the hypoxic ventilatory response, haematology and pectoralis (flight) muscle phenotype of 6 songbird species from 3 families during migratory and non-migratory conditions. Songbirds were captured during southbound migration in southern Ontario, Canada. Half of the birds were assessed during migration, and the rest were transitioned onto a winter photoperiod to induce a non-migratory phenotype and measured. All species exhibited seasonal plasticity at various stages along the O2 cascade, but not all species exhibited the same responses. Songbirds tended to be more hypoxia tolerant during migration, withstanding 5 kPa O2 and breathed more effectively through slower, deeper breaths. Warblers had a stronger haemoglobin-O2 affinity during autumn migration (decrease of ∼4.7 Torr), while the opposite was observed in thrushes (increase of ∼2.6 Torr). In the flight muscle there was an ∼1.2-fold increase in the abundance of muscle fibres with smaller fibre transverse areas during autumn migration, but no changes in capillary:fibre ratio. These adjustments would enhance O2 uptake and transport to the flight muscle. Our findings demonstrate that in the O2 cascade there is no ideal migratory phenotype for all songbirds.
Keywords: Control of breathing; Haematology; Muscle morphology; Thrushes; Vireos; Warblers.
© 2023. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures






References
-
- Bech, C. and Mariussen, C. (2022). Breathing in the cold: seasonal changes in the ventilatory pattern in a small boreal passerine bird. Front. Ecol. Evol. 10, 493. 10.3389/fevo.2022.866102 - DOI
-
- Boggs, D. F., Kilgore, D. L. and Birchard, G. F. (1984). Respiratory physiology of burrowing mammals and birds. Comp. Biochem. Physiol. A Physiol. 77, 1-7. 10.1016/0300-9629(84)90003-3 - DOI
-
- Butler, P. J. (1991). Exercise in birds. J. Exp. Biol. 160, 233-262. 10.1242/jeb.160.1.233 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
