

RUNX1 is required in granulocyte-monocyte progenitors to attenuate inflammatory cytokine production by neutrophils
- PMID: 37536952
- PMCID: PMC10499021
- DOI: 10.1101/gad.350418.123
RUNX1 is required in granulocyte-monocyte progenitors to attenuate inflammatory cytokine production by neutrophils
Abstract
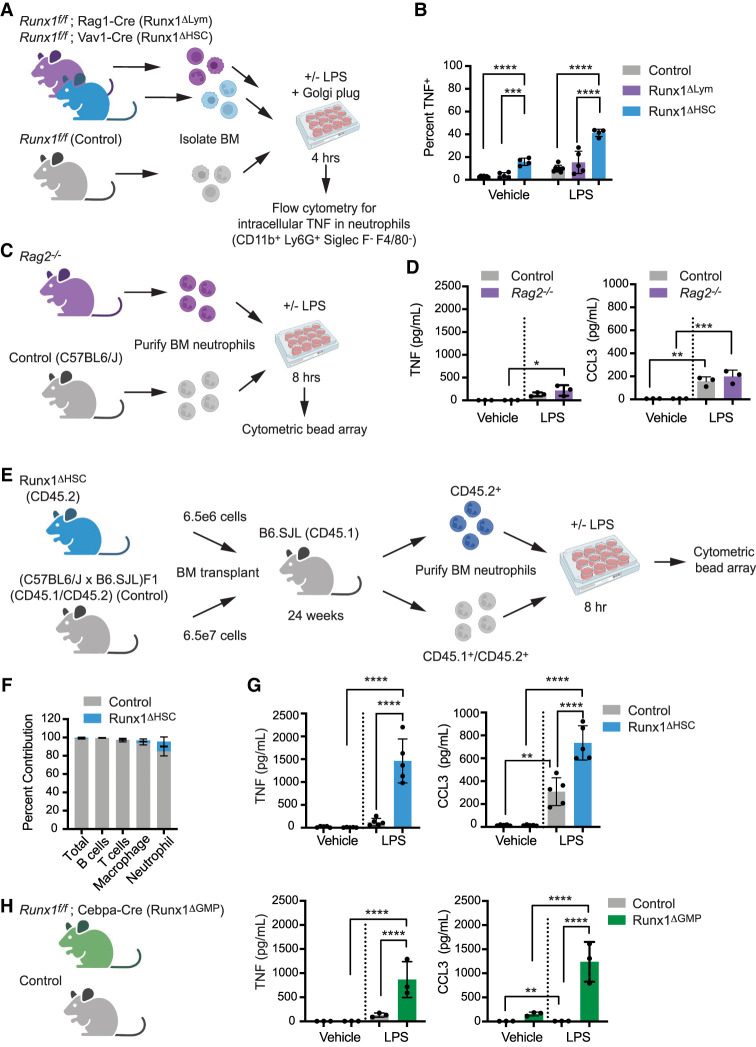
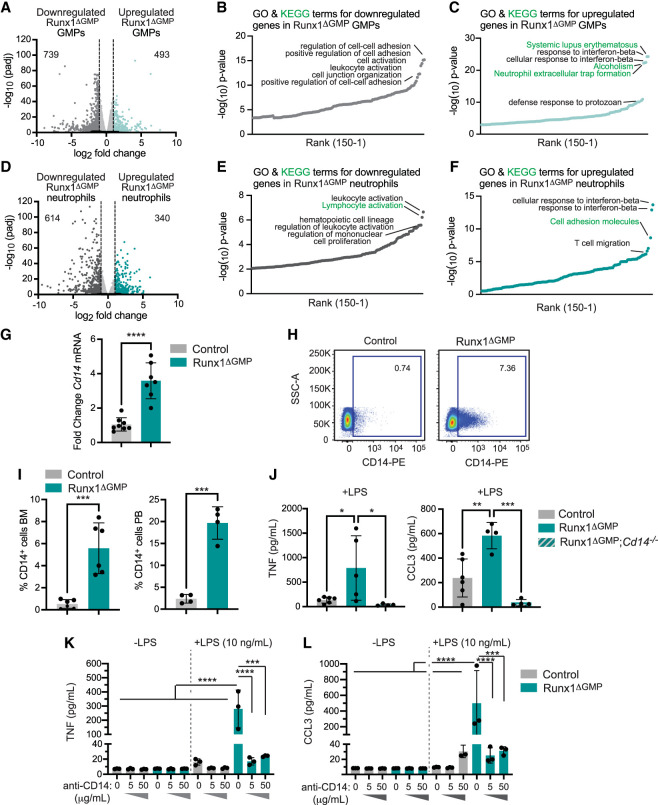
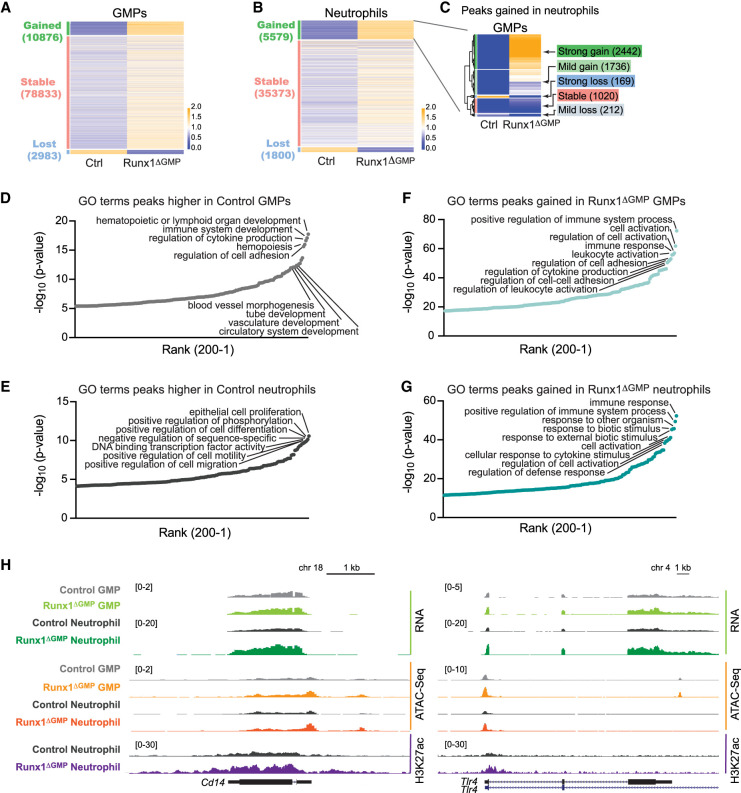
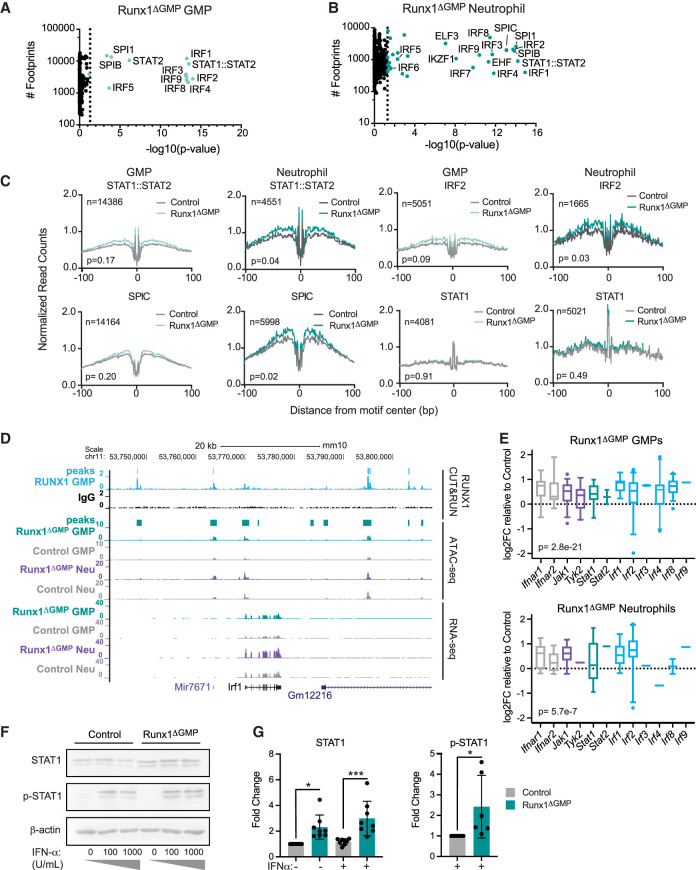
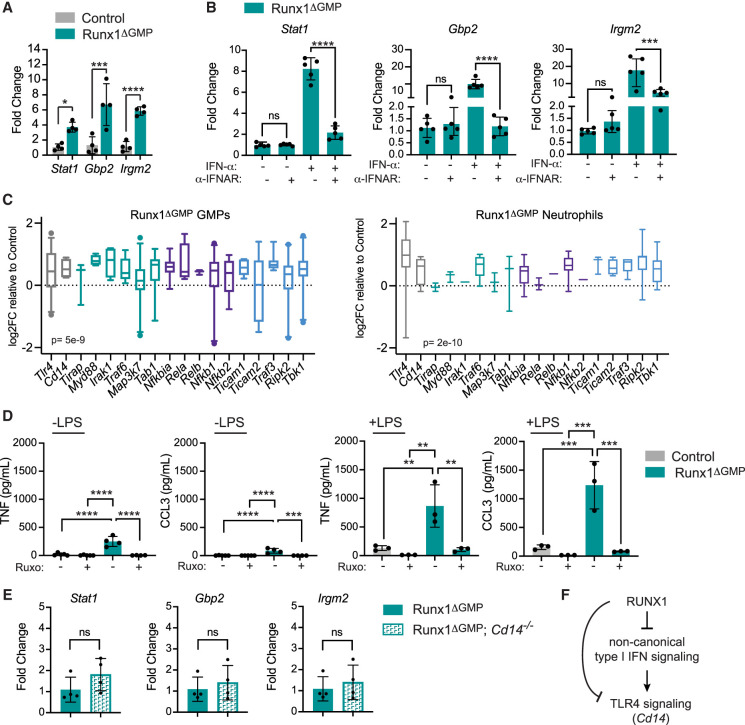
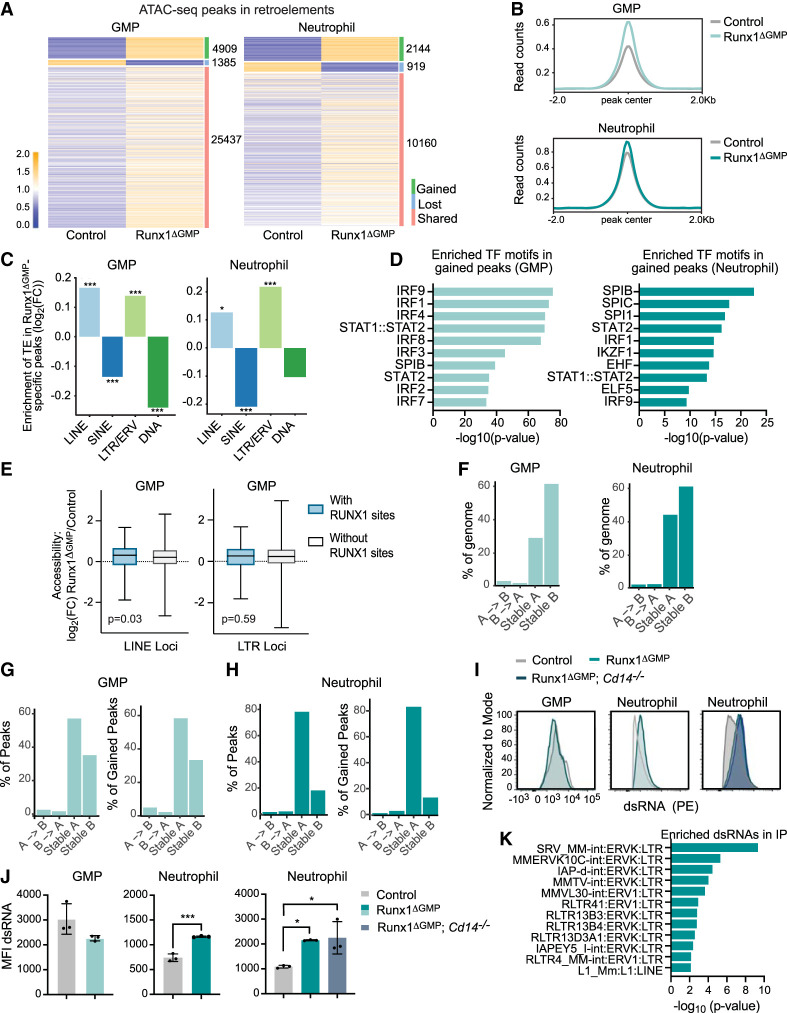
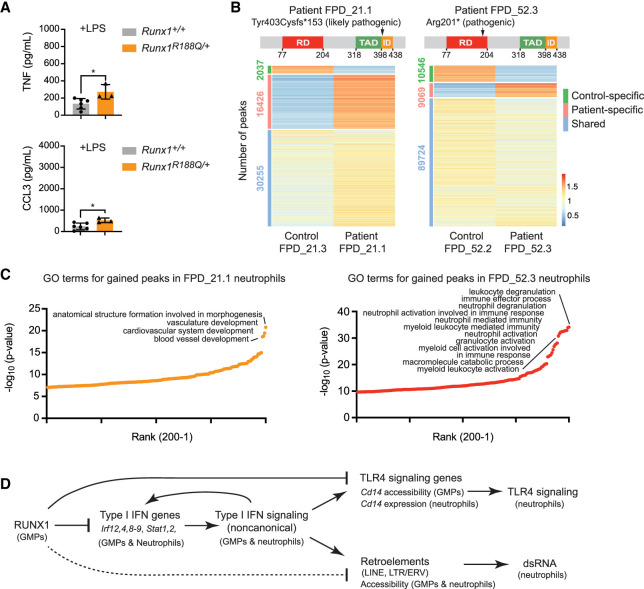
The transcription factor RUNX1 is mutated in familial platelet disorder with associated myeloid malignancy (FPDMM) and in sporadic myelodysplastic syndrome and leukemia. RUNX1 was shown to regulate inflammation in multiple cell types. Here we show that RUNX1 is required in granulocyte-monocyte progenitors (GMPs) to epigenetically repress two inflammatory signaling pathways in neutrophils: Toll-like receptor 4 (TLR4) and type I interferon (IFN) signaling. RUNX1 loss in GMPs augments neutrophils' inflammatory response to the TLR4 ligand lipopolysaccharide through increased expression of the TLR4 coreceptor CD14. RUNX1 binds Cd14 and other genes encoding proteins in the TLR4 and type I IFN signaling pathways whose chromatin accessibility increases when RUNX1 is deleted. Transcription factor footprints for the effectors of type I IFN signaling-the signal transducer and activator of transcription (STAT1::STAT2) and interferon regulatory factors (IRFs)-were enriched in chromatin that gained accessibility in both GMPs and neutrophils when RUNX1 was lost. STAT1::STAT2 and IRF motifs were also enriched in the chromatin of retrotransposons that were derepressed in RUNX1-deficient GMPs and neutrophils. We conclude that a major direct effect of RUNX1 loss in GMPs is the derepression of type I IFN and TLR4 signaling, resulting in a state of fixed maladaptive innate immunity.
Keywords: RUNX1; hematopoiesis; inflammation; retroelements; transposable elements.
© 2023 Zezulin et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Update of
-
RUNX1 is required in granulocyte-monocyte progenitors to attenuate inflammatory cytokine production by neutrophils.bioRxiv [Preprint]. 2023 Jan 28:2023.01.27.525911. doi: 10.1101/2023.01.27.525911. bioRxiv. 2023. Update in: Genes Dev. 2023 Jul 1;37(13-14):605-620. doi: 10.1101/gad.350418.123. PMID: 36747636 Free PMC article. Updated. Preprint.
References
-
- Bellissimo DC, Chen CH, Zhu Q, Bagga S, Lee CT, He B, Wertheim GB, Jordan M, Tan K, Worthen GS, et al. 2020. Runx1 negatively regulates inflammatory cytokine production by neutrophils in response to Toll-like receptor signaling. Blood Adv 4: 1145–1158. 10.1182/bloodadvances.2019000785 - DOI - PMC - PubMed
-
- Brown AL, Arts P, Carmichael CL, Babic M, Dobbins J, Chong CE, Schreiber AW, Feng J, Phillips K, Wang PPS, et al. 2020. RUNX1-mutated families show phenotype heterogeneity and a somatic mutation profile unique to germline predisposed AML. Blood Adv 4: 1131–1144. 10.1182/bloodadvances.2019000901 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous