

Neddylation of phosphoenolpyruvate carboxykinase 1 controls glucose metabolism
- PMID: 37541251
- PMCID: PMC10487638
- DOI: 10.1016/j.cmet.2023.07.003
Neddylation of phosphoenolpyruvate carboxykinase 1 controls glucose metabolism
Abstract
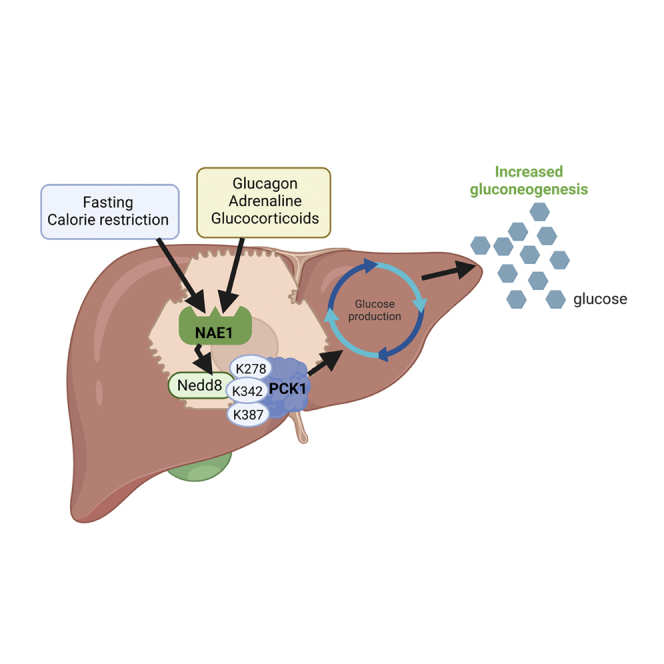
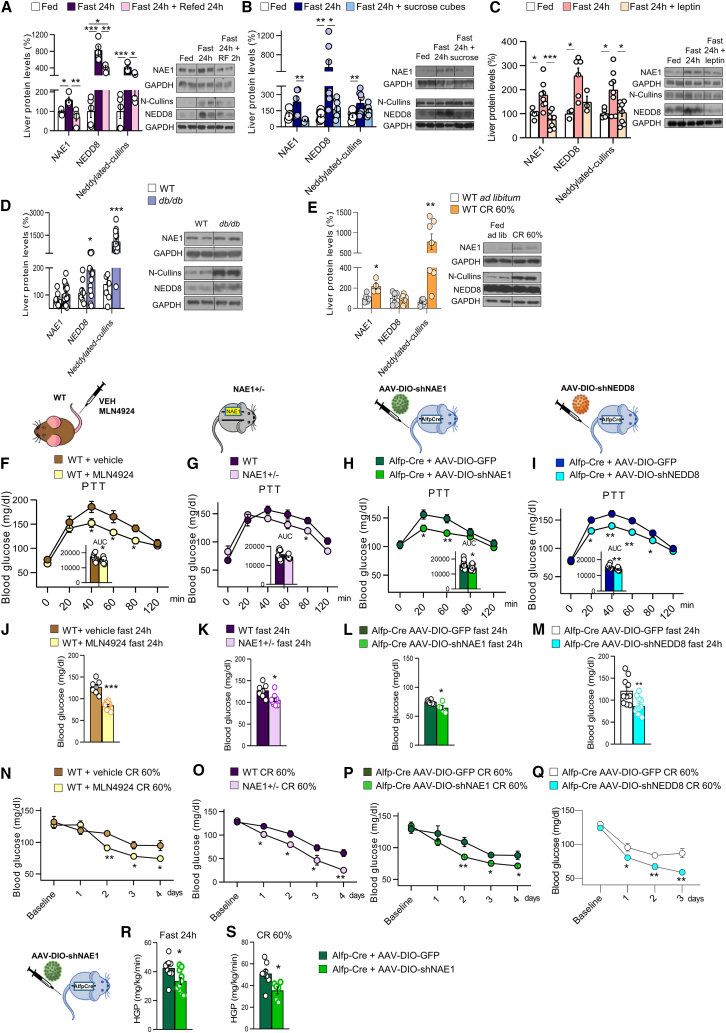
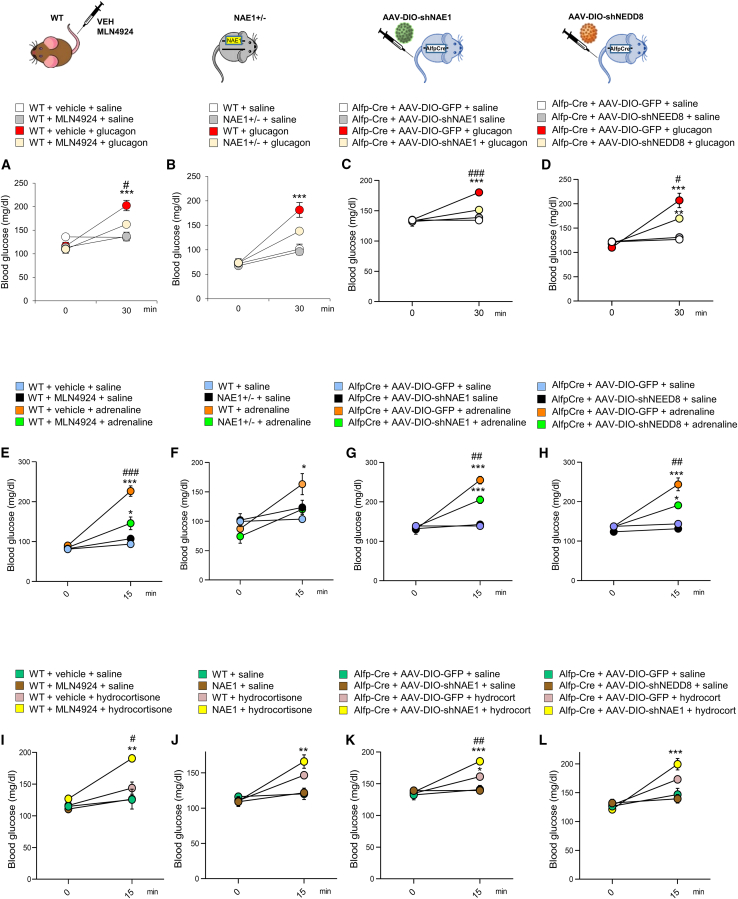
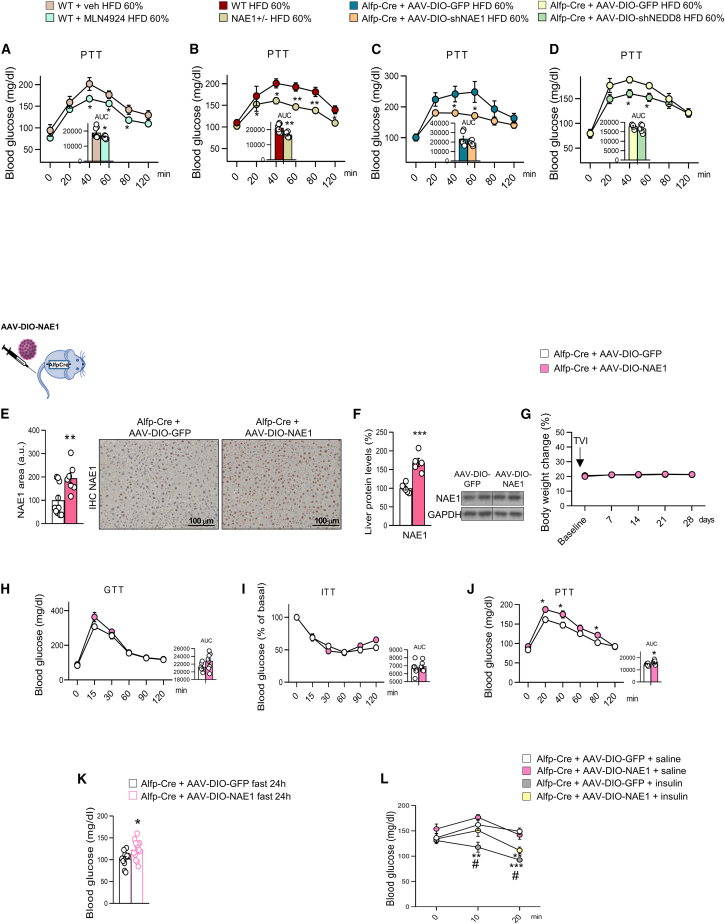
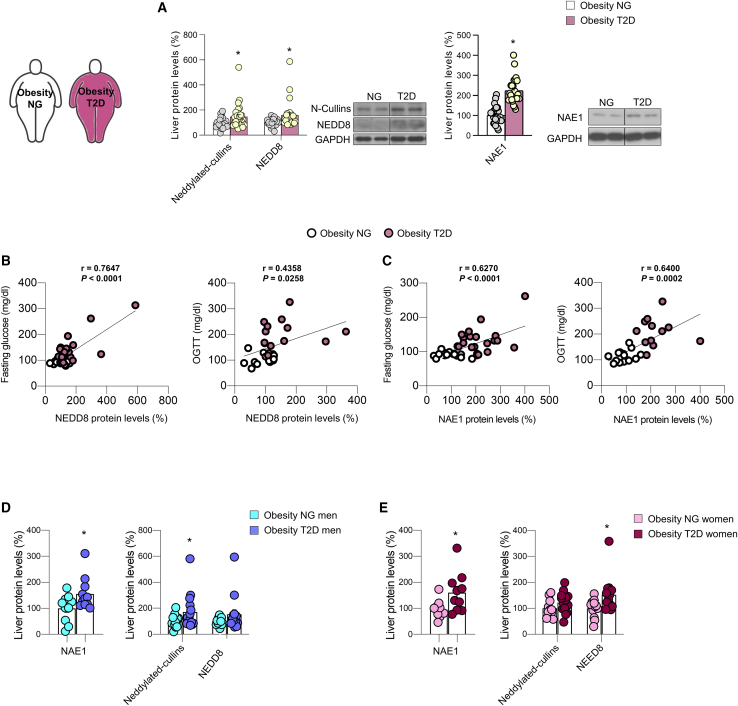
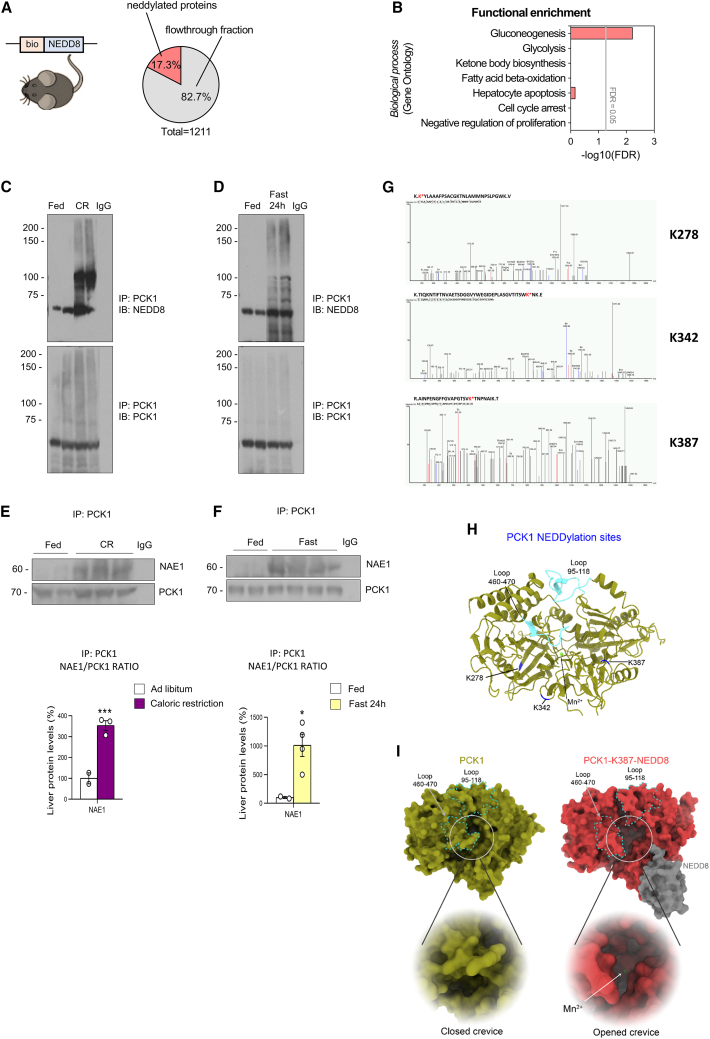
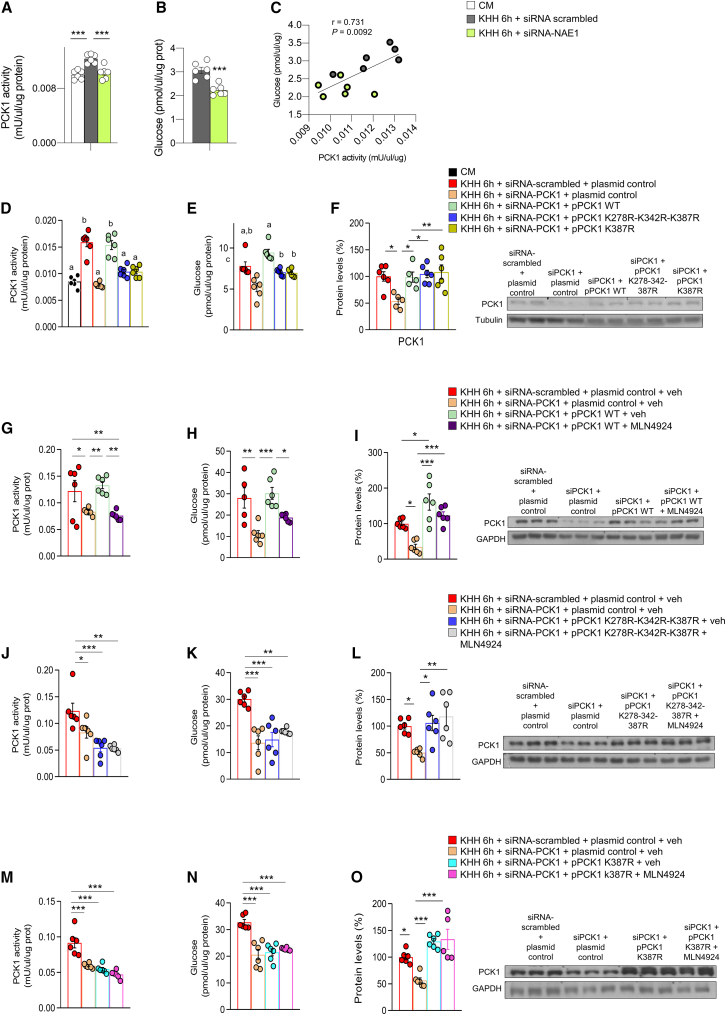
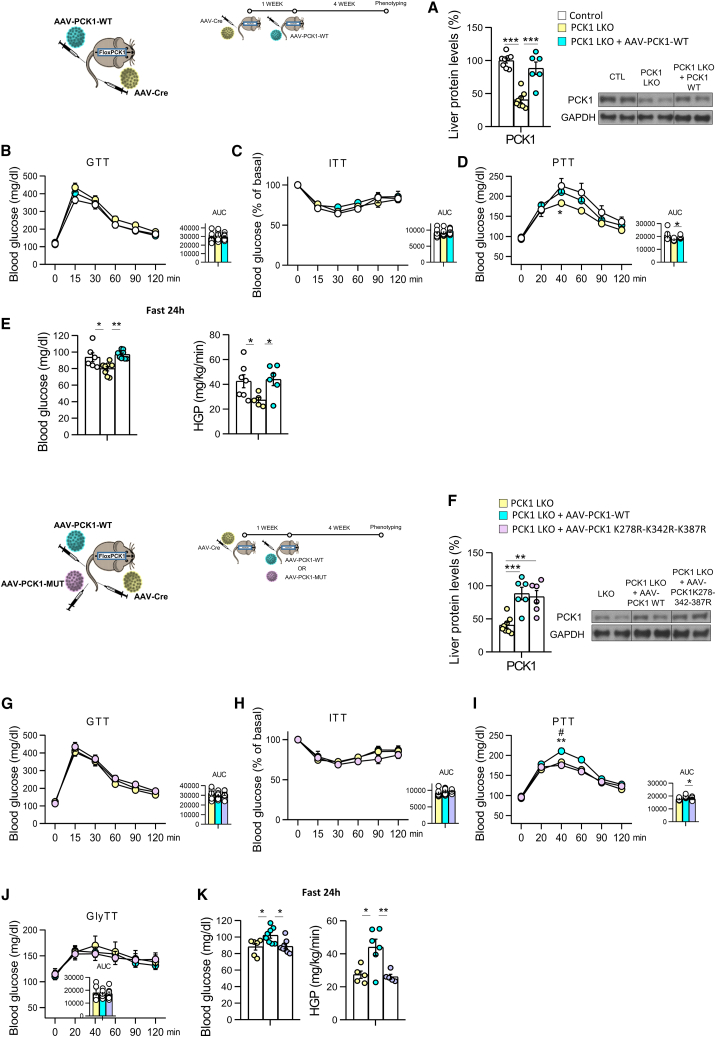
Neddylation is a post-translational mechanism that adds a ubiquitin-like protein, namely neural precursor cell expressed developmentally downregulated protein 8 (NEDD8). Here, we show that neddylation in mouse liver is modulated by nutrient availability. Inhibition of neddylation in mouse liver reduces gluconeogenic capacity and the hyperglycemic actions of counter-regulatory hormones. Furthermore, people with type 2 diabetes display elevated hepatic neddylation levels. Mechanistically, fasting or caloric restriction of mice leads to neddylation of phosphoenolpyruvate carboxykinase 1 (PCK1) at three lysine residues-K278, K342, and K387. We find that mutating the three PCK1 lysines that are neddylated reduces their gluconeogenic activity rate. Molecular dynamics simulations show that neddylation of PCK1 could re-position two loops surrounding the catalytic center into an open configuration, rendering the catalytic center more accessible. Our study reveals that neddylation of PCK1 provides a finely tuned mechanism of controlling glucose metabolism by linking whole nutrient availability to metabolic homeostasis.
Keywords: PCK1; calorie restriction; fasting; glucagon; glucose metabolism; neddylation; type 2 diabetes.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
