

Enolase represents a metabolic checkpoint controlling the differential exhaustion programmes of hepatitis virus-specific CD8+ T cells
- PMID: 37541771
- PMCID: PMC10511960
- DOI: 10.1136/gutjnl-2022-328734
Enolase represents a metabolic checkpoint controlling the differential exhaustion programmes of hepatitis virus-specific CD8+ T cells
Abstract
Objective: Exhausted T cells with limited effector function are enriched in chronic hepatitis B and C virus (HBV and HCV) infection. Metabolic regulation contributes to exhaustion, but it remains unclear how metabolism relates to different exhaustion states, is impacted by antiviral therapy, and if metabolic checkpoints regulate dysfunction.
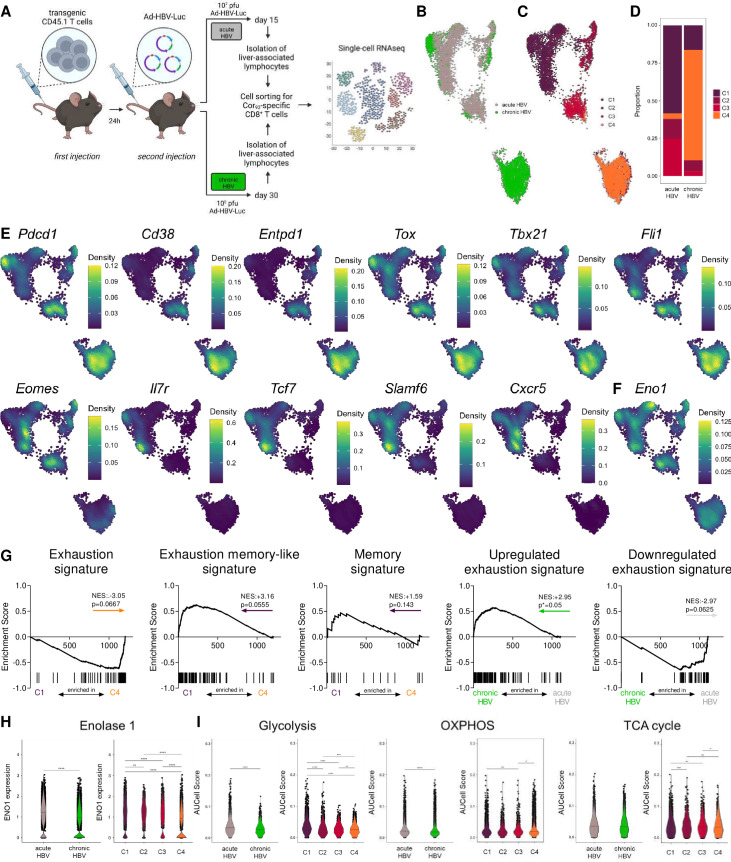
Design: Metabolic state, exhaustion and transcriptome of virus-specific CD8+ T cells from chronic HBV-infected (n=31) and HCV-infected patients (n=52) were determined ex vivo and during direct-acting antiviral (DAA) therapy. Metabolic flux and metabolic checkpoints were tested in vitro. Intrahepatic virus-specific CD8+ T cells were analysed by scRNA-Seq in a HBV-replicating murine in vivo model of acute and chronic infection.
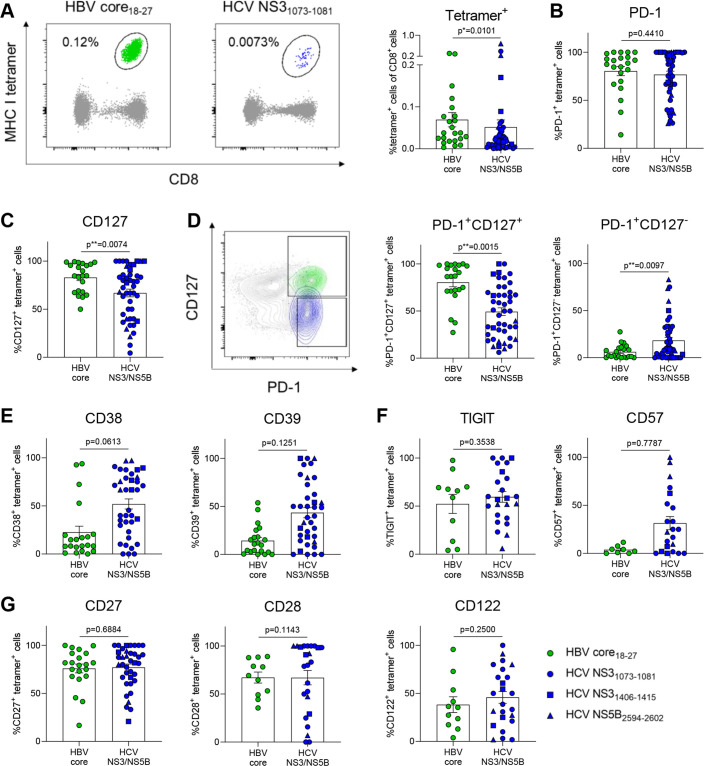
Results: HBV-specific (core18-27, polymerase455-463) and HCV-specific (NS31073-1081, NS31406-1415, NS5B2594-2602) CD8+ T cell responses exhibit heterogeneous metabolic profiles connected to their exhaustion states. The metabolic state was connected to the exhaustion profile rather than the aetiology of infection. Mitochondrial impairment despite intact glucose uptake was prominent in severely exhausted T cells linked to elevated liver inflammation in chronic HCV infection and in HBV polymerase455-463 -specific CD8+ T cell responses. In contrast, relative metabolic fitness was observed in HBeAg-negative HBV infection in HBV core18-27-specific responses. DAA therapy partially improved mitochondrial programmes in severely exhausted HCV-specific T cells and enriched metabolically fit precursors. We identified enolase as a metabolic checkpoint in exhausted T cells. Metabolic bypassing improved glycolysis and T cell effector function. Similarly, enolase deficiency was observed in intrahepatic HBV-specific CD8+ T cells in a murine model of chronic infection.
Conclusion: Metabolism of HBV-specific and HCV-specific T cells is strongly connected to their exhaustion severity. Our results highlight enolase as metabolic regulator of severely exhausted T cells. They connect differential bioenergetic fitness with distinct exhaustion subtypes and varying liver disease, with implications for therapeutic strategies.
Keywords: alpha beta T cells; chronic viral hepatitis; hepatitis B; hepatitis C; immunology in hepatology.
© Author(s) (or their employer(s)) 2023. Re-use permitted under CC BY. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures





Comment in
-
Enolase: a metabolic checkpoint behind diverse exhaustion stages of CD8+ T cells in chronic HBV and HCV.Gut. 2023 Oct;72(10):1814-1815. doi: 10.1136/gutjnl-2023-330541. Epub 2023 Sep 6. Gut. 2023. PMID: 37673656 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials