

Palmitoyltransferase ZDHHC3 Aggravates Nonalcoholic Steatohepatitis by Targeting S-Palmitoylated IRHOM2
- PMID: 37544908
- PMCID: PMC10558657
- DOI: 10.1002/advs.202302130
Palmitoyltransferase ZDHHC3 Aggravates Nonalcoholic Steatohepatitis by Targeting S-Palmitoylated IRHOM2
Erratum in
-
Palmitoyltransferase ZDHHC3 Aggravates Nonalcoholic Steatohepatitis by Targeting S-Palmitoylated IRHOM2.Adv Sci (Weinh). 2023 Dec;10(36):e2308024. doi: 10.1002/advs.202308024. Adv Sci (Weinh). 2023. PMID: 38153358 Free PMC article. No abstract available.
Abstract
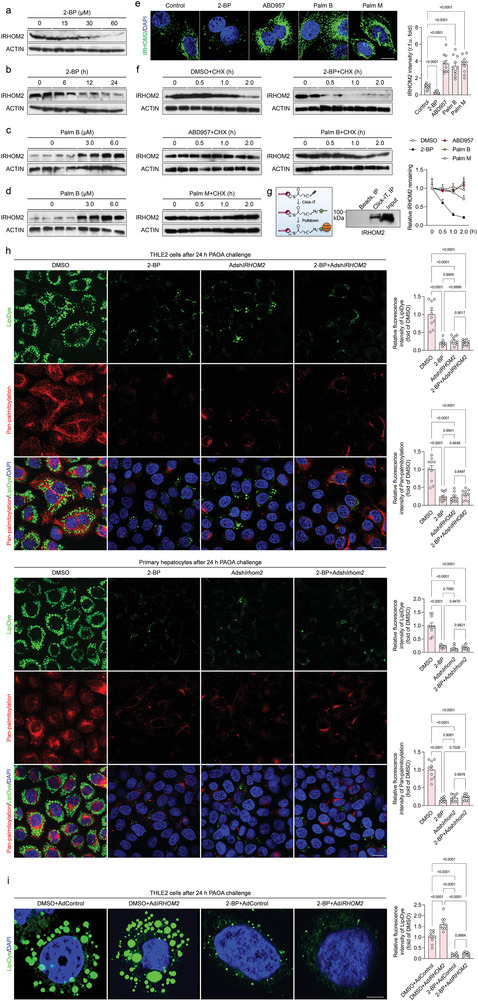
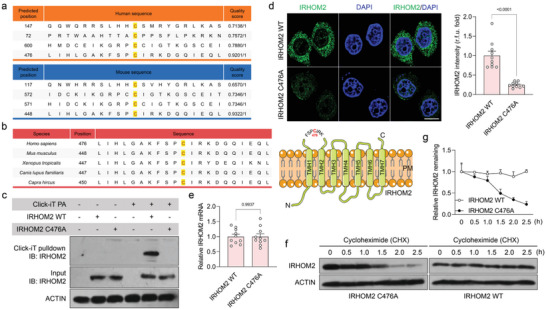
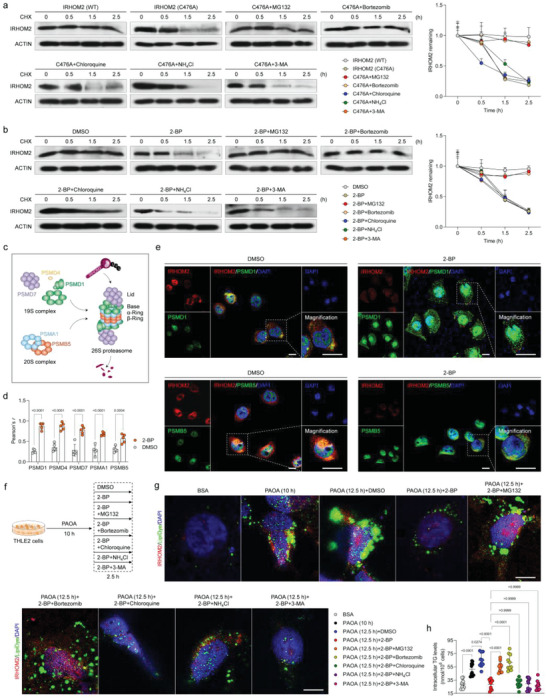
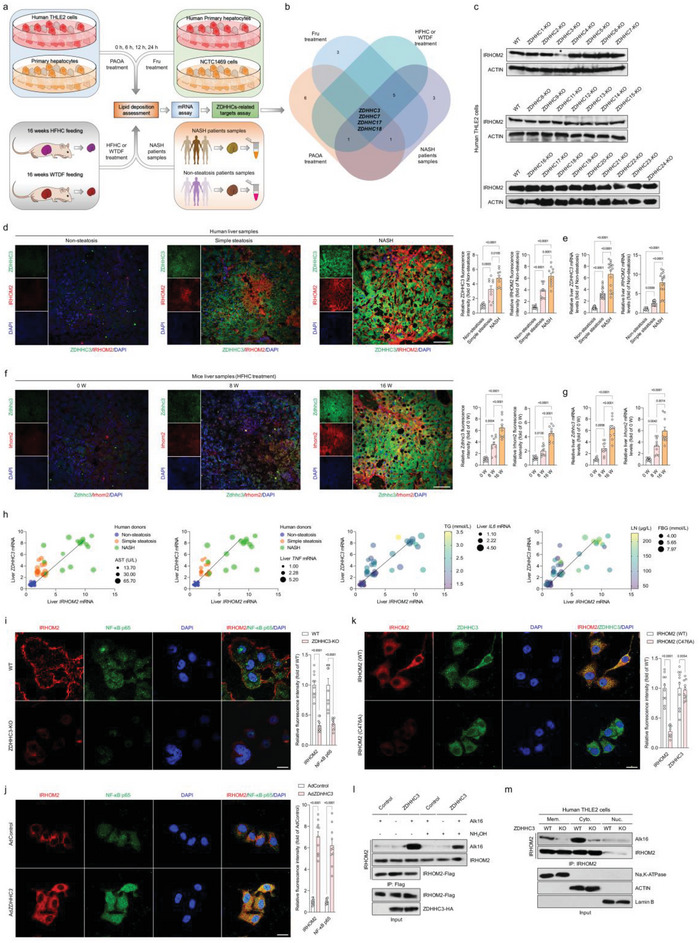
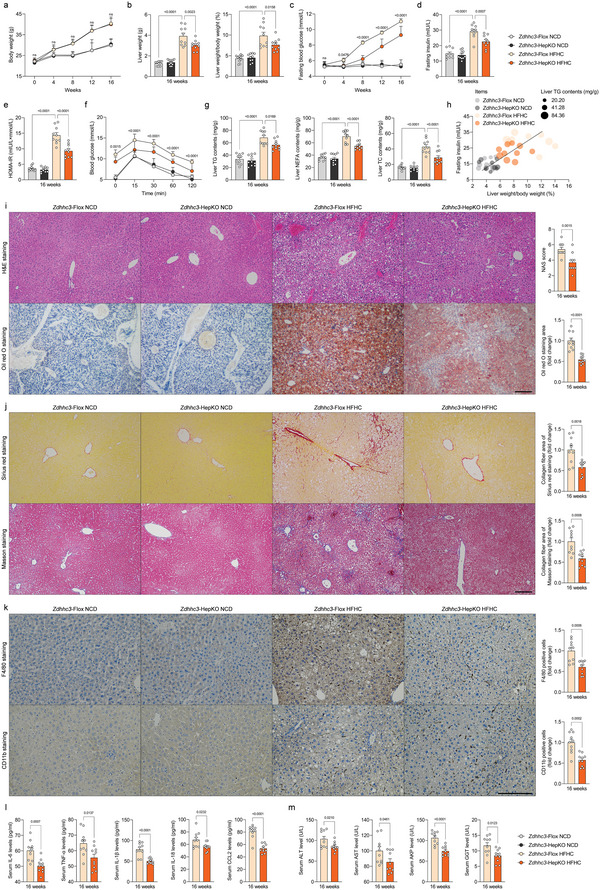
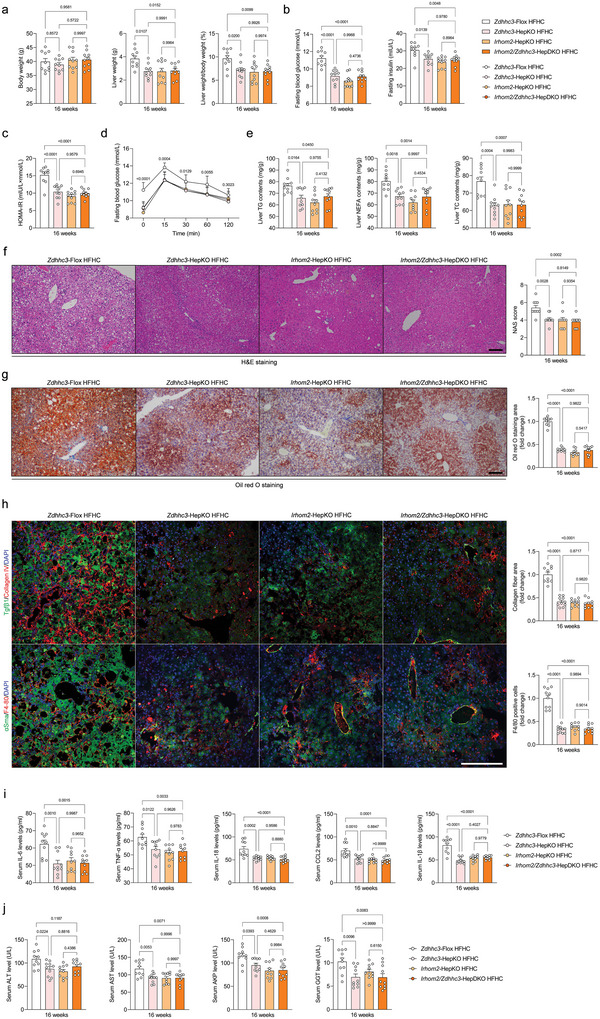
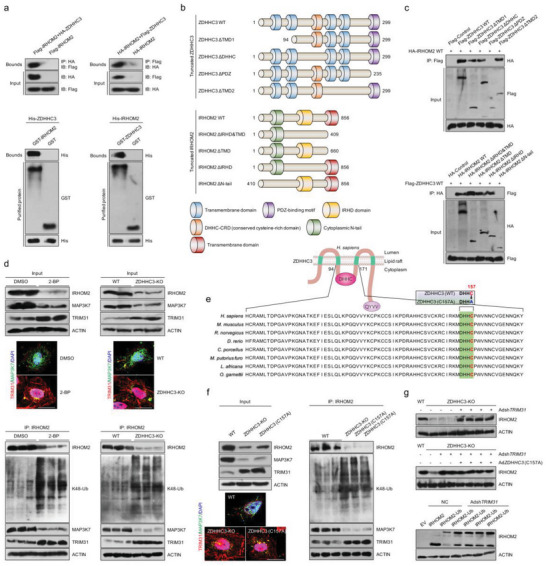
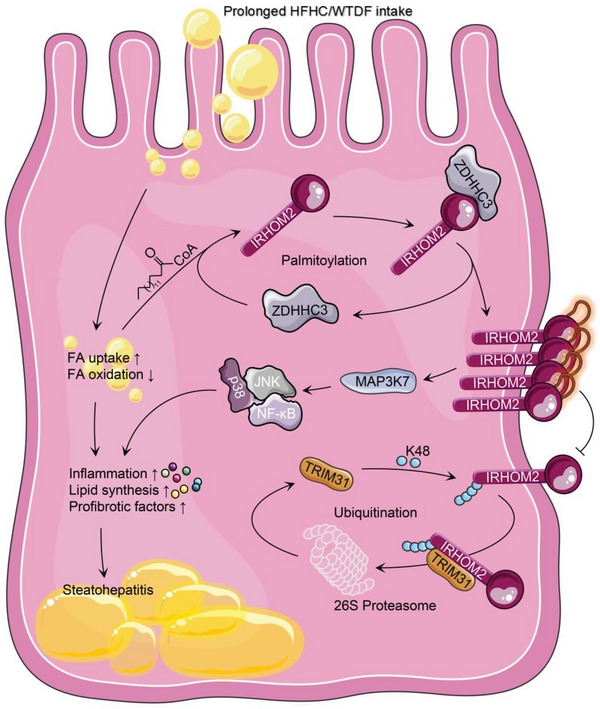
Underestimation of the complexity of pathogenesis in nonalcoholic steatohepatitis (NASH) significantly encumbers development of new drugs and targeted therapy strategies. Inactive rhomboid protein 2 (IRHOM2) has a multifunctional role in regulating inflammation, cell survival, and immunoreaction. Although cytokines and chemokines promote IRHOM2 trafficking or cooperate with partner factors by phosphorylation or ubiquitin ligases-mediated ubiquitination to perform physiological process, it remains unknown whether other regulators induce IRHOM2 activation via different mechanisms in NASH progression. Here the authors find that IRHOM2 is post-translationally S-palmitoylated at C476 in iRhom homology domain (IRHD), which facilitates its cytomembrane translocation and stabilization. Fatty-acids challenge can directly promote IRHOM2 trafficking by increasing its palmitoylation. Additionally, the authors identify Zinc finger DHHC-type palmitoyltransferase 3 (ZDHHC3) as a key acetyltransferase required for the IRHOM2 palmitoylation. Fatty-acids administration enhances IRHOM2 palmitoylation by increasing the direct association between ZDHHC3 and IRHOM2, which is catalyzed by the DHHC (C157) domain of ZDHHC3. Meanwhile, a metabolic stresses-triggered increase of ZDHHC3 maintains palmitoylated IRHOM2 accumulation by blocking its ubiquitination, consequently suppressing its ubiquitin-proteasome-related degradation mediated by tripartite motif containing 31 (TRIM31). High-levels of ZDHHC3 protein abundance positively correlate with the severity of NASH phenotype in patient samples. Hepatocyte-specific dysfunction of ZDHHC3 significantly inhibits palmitoylated IRHOM2 deposition, therefore suppressing the fatty-acids-mediated hepatosteatosis and inflammation in vitro, as well as NASH pathological phenotype induced by two different high-energy diets (HFHC & WTDF) in the in vivo rodent and rabbit model. Inversely, specific restoration of ZDHHC3 in hepatocytes markedly provides acceleration over the course of NASH development via increasing palmitoylation of IRHOM2 along with suppression of ubiquitin degradation. The current work uncovers that ZDHHC3-induced palmitoylation is a novel regulatory mechanism and signal that regulates IRHOM2 trafficking, which confers evidence associating the regulation of palmitoylation with NASH progression.
Keywords: Palmitoylation; hepatosteatosis; inactive rhomboid protein 2 (IRHOM2); nonalcoholic steatohepatitis (NASH); tripartite motif containing 31 (TRIM31); zinc finger DHHC-type palmitoyltransferase 3 (ZDHHC3).
© 2023 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Quek J., Chan K. E., Wong Z. Y., Tan C., Tan B., Lim W. H., Tan Darren J. H., Tang Ansel S. P., Tay P., Xiao J., Yong J. N., Zeng R. W., Chew Nicholas W. S., Nah B., Kulkarni A., Siddiqui M. S., Dan Y. Y., Wong Vincent W.‐S., Sanyal A. J., Noureddin M., Muthiah M., Ng C. H., Lancet Gastroenterol Hepatol 2023, 8, 20. - PubMed
-
- Eguchi Y., Wong G., Lee I.‐H., Akhtar O., Lopes R., Sumida Y., Hepatol. Res. 2021, 51, 19. - PubMed
-
- Anstee Q. M., Reeves H. L., Kotsiliti E., Govaere O., Heikenwalder M., Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 411. - PubMed
-
- Cotter T. G., Rinellaa M., Gastroenterology 2020, 158, 1851. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 82200652/National Natural Science Foundation of China
- cstc2017jcyjAX0356/Chongqing Research Program of Basic Research and Frontier Technology
- cstc2018jcyjA3686/Chongqing Research Program of Basic Research and Frontier Technology
- cstc2018jcyjAX0784/Chongqing Research Program of Basic Research and Frontier Technology
- cstc2018jcyjA1472/Chongqing Research Program of Basic Research and Frontier Technology
- cstc2018jcyjAX0811/Chongqing Research Program of Basic Research and Frontier Technology
- cstc2018jcyjA3533/Chongqing Research Program of Basic Research and Frontier Technology
- KJZD-M201801601/Chongqing Research Program of Basic Research and Frontier Technology
- KY201710B/Chongqing Research Program of Basic Research and Frontier Technology
- 17GZKP01/Chongqing Research Program of Basic Research and Frontier Technology
- KJQN201901608/Science and Technology Research Program of Chongqing Education Commission of China
- KJQN201901615/Science and Technology Research Program of Chongqing Education Commission of China
- KJ1601402/Science and Technology Research Program of Chongqing Education Commission of China
- CSDP19FSO1108/Science and Technology Research Program of Chongqing Education Commission of China
- KJZD-K202001603/Science and Technology Research Program of Chongqing Education Commission of China
- CSTB2022NSCQ-LZX0053/Natural Science Foundation of Chongqing, China
- CSTB2022NSCQ-MSX0947/Natural Science Foundation of Chongqing, China
- CSTB2022NSCQ-MSX5364/Natural Science Foundation of Chongqing, China
- CQCY201903258cstc2021ycjh-bgzxm0202/Chongqing Professional Talents Plan for Innovation and Entrepreneurship Demonstration Team
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases