

Branchpoints as potential targets of exon-skipping therapies for genetic disorders
- PMID: 37547287
- PMCID: PMC10403725
- DOI: 10.1016/j.omtn.2023.07.011
Branchpoints as potential targets of exon-skipping therapies for genetic disorders
Abstract
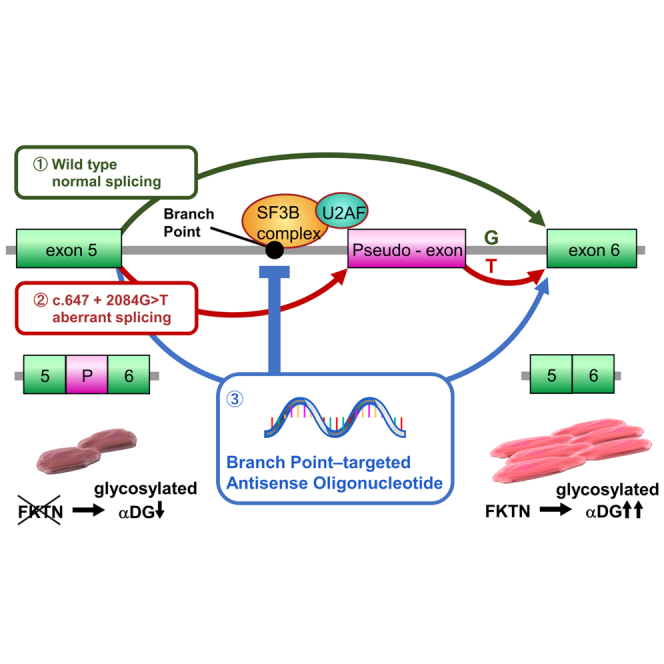
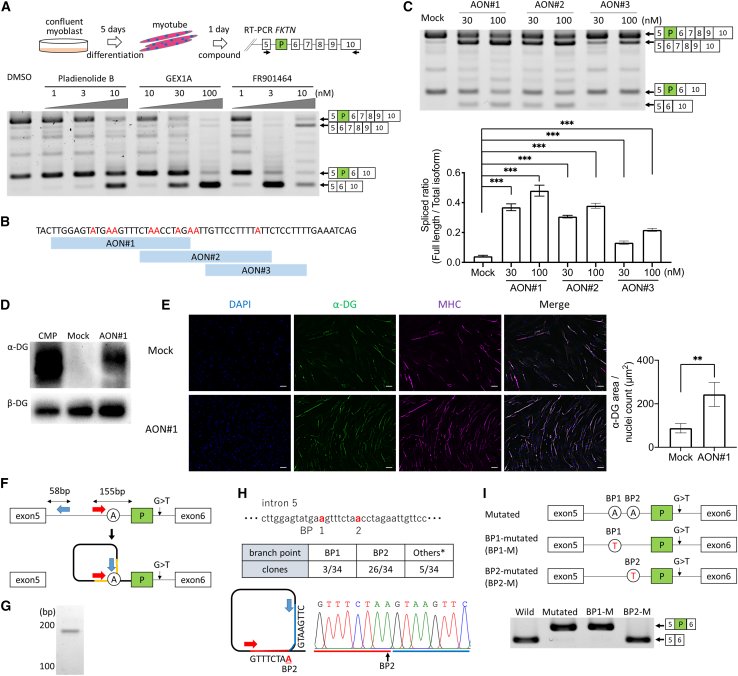
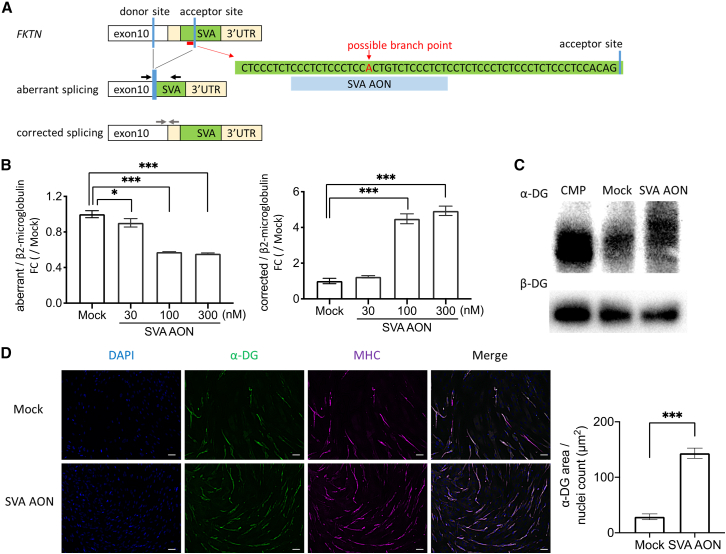
Fukutin (FKTN) c.647+2084G>T creates a pseudo-exon with a premature stop codon, which causes Fukuyama congenital muscular dystrophy (FCMD). We aimed to ameliorate aberrant splicing of FKTN caused by this variant. We screened compounds focusing on splicing regulation using the c.647+2084G>T splicing reporter and discovered that the branchpoint, which is essential for splicing reactions, could be a potential therapeutic target. To confirm the effectiveness of branchpoints as targets for exon skipping, we designed branchpoint-targeted antisense oligonucleotides (BP-AONs). This restored normal FKTN mRNA and protein production in FCMD patient myotubes. We identified a functional BP by detecting splicing intermediates and creating BP mutations in the FKTN reporter gene; this BP was non-redundant and sufficiently blocked by BP-AONs. Next, a BP-AON was designed for a different FCMD-causing variant, which induces pathogenic exon trapping by a common SINE-VNTR-Alu-type retrotransposon. Notably, this BP-AON also restored normal FKTN mRNA and protein production in FCMD patient myotubes. Our findings suggest that BPs could be potential targets in exon-skipping therapeutic strategies for genetic disorders.
Keywords: MT: Oligonucleotides: Therapies and Applications; antisense oligonucleotide; branchpoint; exon-skipping therapy; muscular dystrophy; splice-switching; splicing regulatory elements.
© 2023 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Vaz-Drago R., Custódio N., Carmo-Fonseca M. Deep intronic mutations and human disease. Hum. Genet. 2017;136:1093–1111. - PubMed
-
- Okubo M., Noguchi S., Awaya T., Hosokawa M., Tsukui N., Ogawa M., Hayashi S., Komaki H., Mori-Yoshimura M., Oya Y., et al. RNA-seq analysis, targeted long-read sequencing and in silico prediction to unravel pathogenic intronic events and complicated splicing abnormalities in dystrophinopathy. Hum. Genet. 2023;142:59–71. - PubMed
-
- Fukuyama Y., Osawa M., Suzuki H. Congenital progressive muscular dystrophy of the Fukuyama type - clinical, genetic and pathological considerations. Brain Dev. 1981;3:1–29. - PubMed
-
- Kanagawa M., Kobayashi K., Tajiri M., Manya H., Kuga A., Yamaguchi Y., Akasaka-Manya K., Furukawa J.I., Mizuno M., Kawakami H., et al. Identification of a Post-translational Modification with Ribitol-Phosphate and Its Defect in Muscular Dystrophy. Cell Rep. 2016;14:2209–2223. - PubMed
-
- Muntoni F., Brockington M., Blake D.J., Torelli S., Brown S.C. Defective glycosylation in muscular dystrophy. Lancet. 2002;360:1419–1421. - PubMed
