

Functional unknomics: Systematic screening of conserved genes of unknown function
- PMID: 37552676
- PMCID: PMC10409296
- DOI: 10.1371/journal.pbio.3002222
Functional unknomics: Systematic screening of conserved genes of unknown function
Abstract
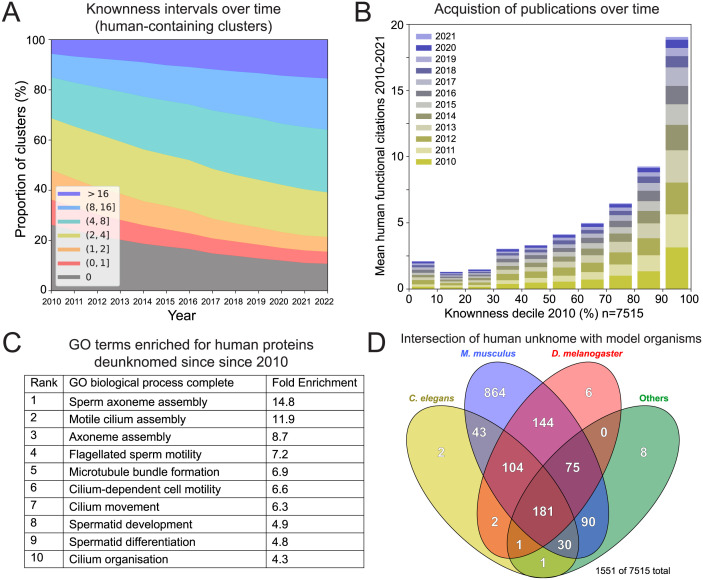
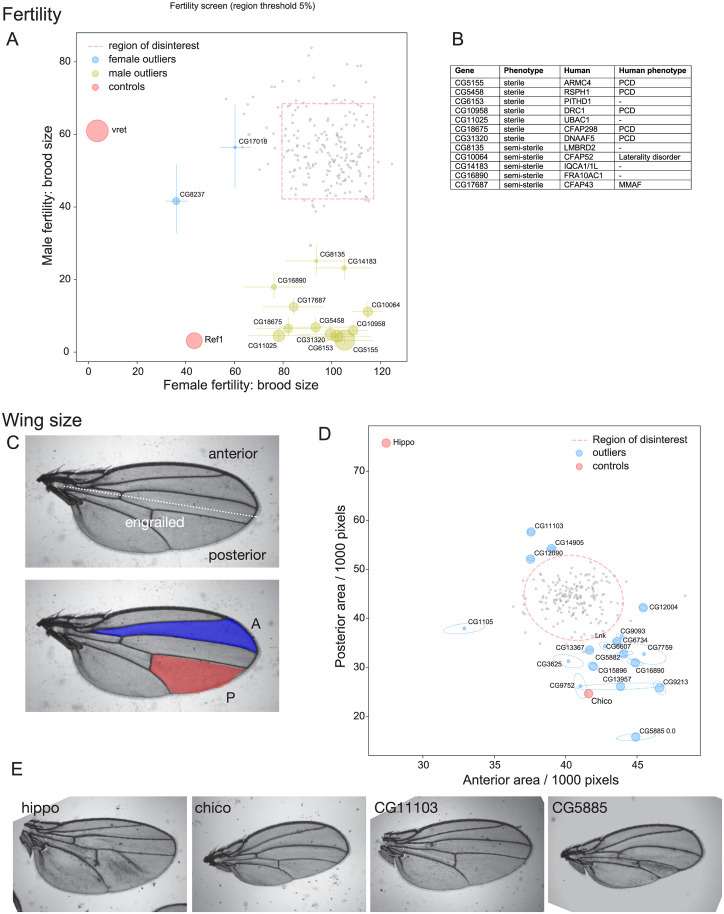
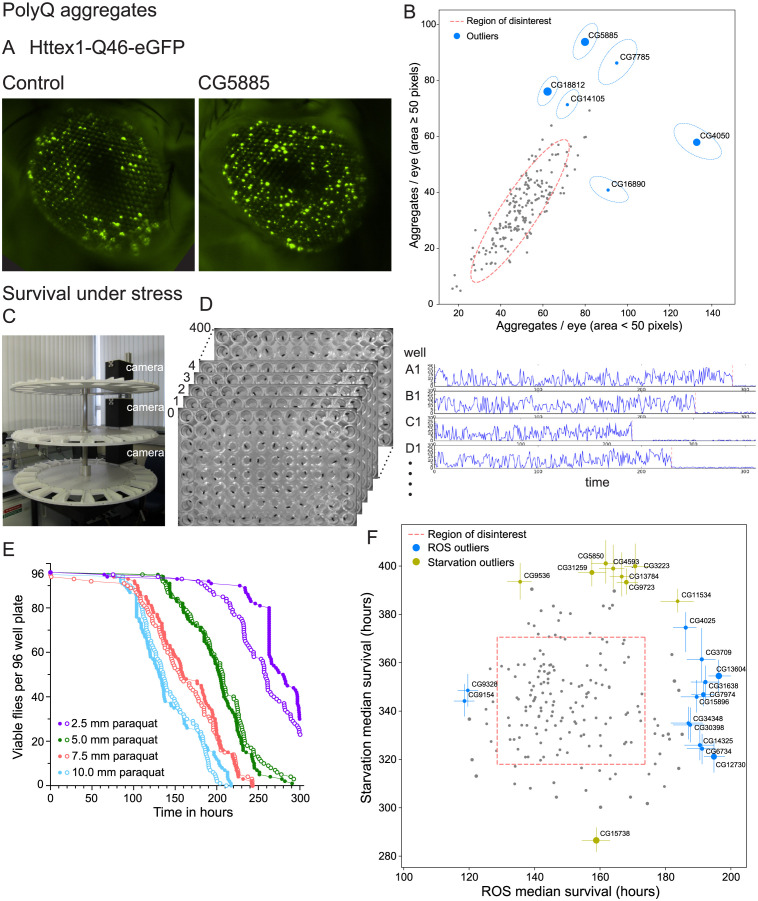
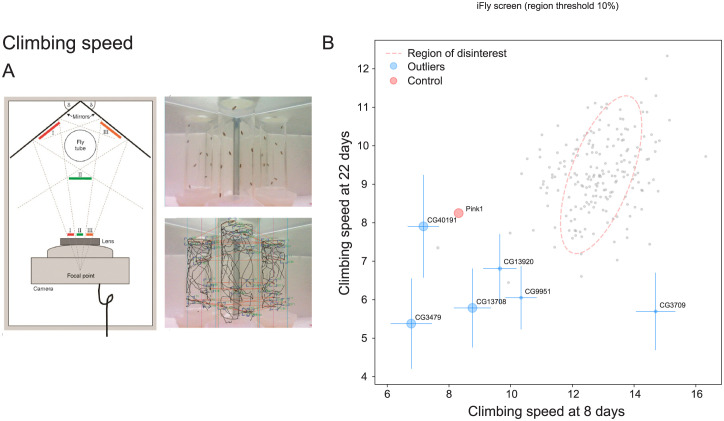
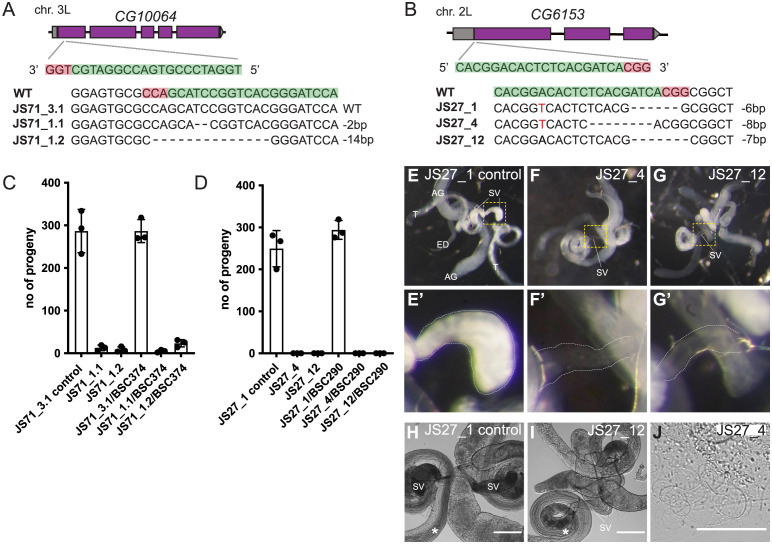
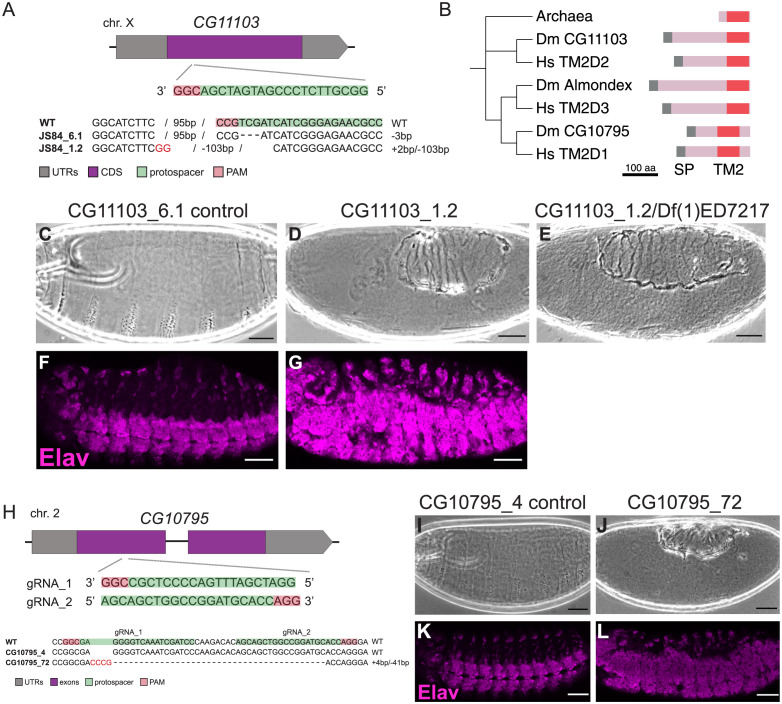
The human genome encodes approximately 20,000 proteins, many still uncharacterised. It has become clear that scientific research tends to focus on well-studied proteins, leading to a concern that poorly understood genes are unjustifiably neglected. To address this, we have developed a publicly available and customisable "Unknome database" that ranks proteins based on how little is known about them. We applied RNA interference (RNAi) in Drosophila to 260 unknown genes that are conserved between flies and humans. Knockdown of some genes resulted in loss of viability, and functional screening of the rest revealed hits for fertility, development, locomotion, protein quality control, and resilience to stress. CRISPR/Cas9 gene disruption validated a component of Notch signalling and 2 genes contributing to male fertility. Our work illustrates the importance of poorly understood genes, provides a resource to accelerate future research, and highlights a need to support database curation to ensure that misannotation does not erode our awareness of our own ignorance.
Copyright: © 2023 Rocha et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Sinha S, Eisenhaber B, Jensen LJ, Kalbuaji B, Eisenhaber F. Darkness in the human gene and protein function space: widely modest or absent illumination by the life science literature and the trend for fewer protein function discoveries since 2000. Proteomics. 2018;18:e1800093. doi: 10.1002/pmic.201800093 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
