

Natural selection has driven the recurrent loss of an immunity gene that protects Drosophila against a major natural parasite
- PMID: 37552757
- PMCID: PMC10438844
- DOI: 10.1073/pnas.2211019120
Natural selection has driven the recurrent loss of an immunity gene that protects Drosophila against a major natural parasite
Abstract
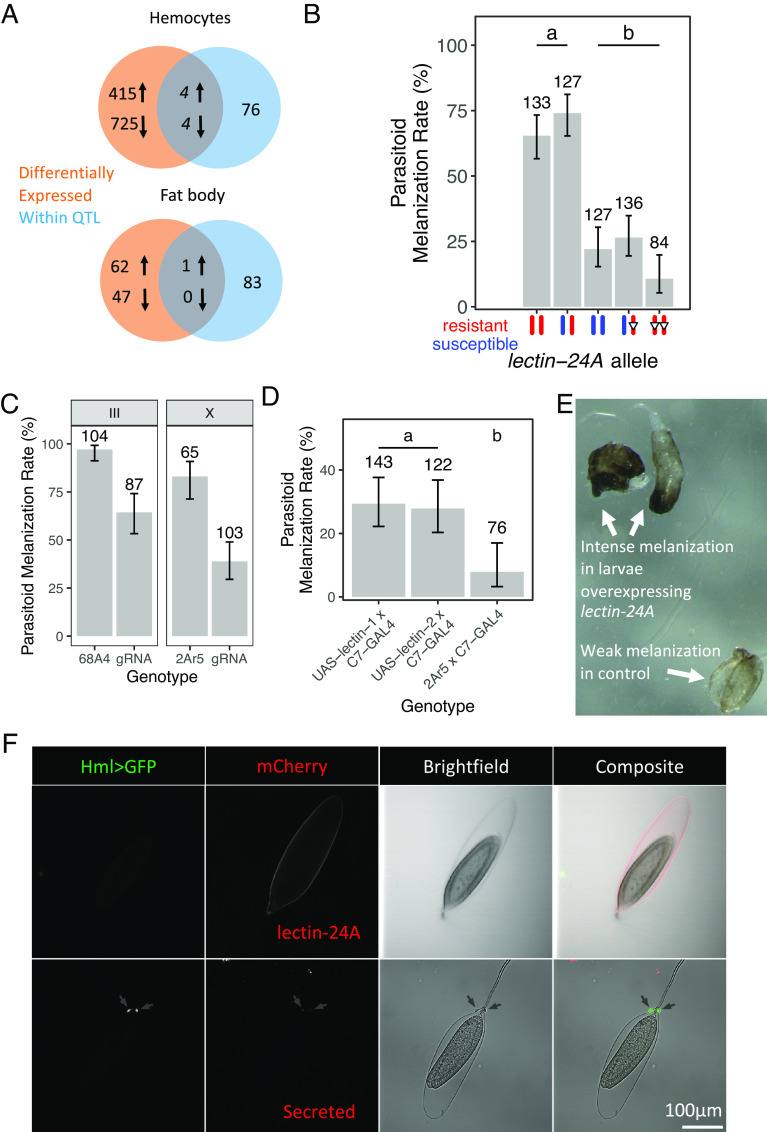
Polymorphisms in immunity genes can have large effects on susceptibility to infection. To understand the origins of this variation, we have investigated the genetic basis of resistance to the parasitoid wasp Leptopilina boulardi in Drosophila melanogaster. We found that increased expression of the gene lectin-24A after infection by parasitic wasps was associated with a faster cellular immune response and greatly increased rates of killing the parasite. lectin-24A encodes a protein that is strongly up-regulated in the fat body after infection and localizes to the surface of the parasite egg. In certain susceptible lines, a deletion upstream of the lectin-24A has largely abolished expression. Other mutations predicted to abolish the function of this gene have arisen recurrently in this gene, with multiple loss-of-expression alleles and premature stop codons segregating in natural populations. The frequency of these alleles varies greatly geographically, and in some southern African populations, natural selection has driven them near to fixation. We conclude that natural selection has favored the repeated loss of an important component of the immune system, suggesting that in some populations, a pleiotropic cost to lectin-24A expression outweighs the benefits of resistance.
Keywords: C-type lectin; Leptopilina boulardi; cis-regulatory polymorphism; loss of function; melanization.
Conflict of interest statement
The authors declare no competing interest.
Figures





Similar articles
-
Adaptive evolution of a novel Drosophila lectin induced by parasitic wasp attack.Mol Biol Evol. 2012 Feb;29(2):565-77. doi: 10.1093/molbev/msr191. Epub 2011 Aug 26. Mol Biol Evol. 2012. PMID: 21873297 Free PMC article.
-
An introduction to parasitic wasps of Drosophila and the antiparasite immune response.J Vis Exp. 2012 May 7;(63):e3347. doi: 10.3791/3347. J Vis Exp. 2012. PMID: 22588641 Free PMC article.
-
Variation of Leptopilina boulardi success in Drosophila hosts: what is inside the black box?Adv Parasitol. 2009;70:147-88. doi: 10.1016/S0065-308X(09)70006-5. Adv Parasitol. 2009. PMID: 19773070 Review.
-
Genetic interactions between the parasitoid wasp Leptopilina boulardi and its Drosophila hosts.Heredity (Edinb). 2007 Jan;98(1):21-7. doi: 10.1038/sj.hdy.6800893. Epub 2006 Oct 11. Heredity (Edinb). 2007. PMID: 17035953
-
Immunogenetic aspects of the cellular immune response of Drosophilia against parasitoids.Immunogenetics. 2001;52(3-4):157-64. doi: 10.1007/s002510000272. Immunogenetics. 2001. PMID: 11220617 Review.
Cited by
-
Encapsulation and Melanization Are Not Correlated to Successful Immune Defense Against Parasitoid Wasps in Drosophila melanogaster.Cells. 2025 Jan 3;14(1):46. doi: 10.3390/cells14010046. Cells. 2025. PMID: 39791747 Free PMC article.
-
Experimental horizontal transfer of phage-derived genes to Drosophila confers innate immunity to parasitoids.Curr Biol. 2025 Feb 3;35(3):514-529.e7. doi: 10.1016/j.cub.2024.11.071. Epub 2024 Dec 20. Curr Biol. 2025. PMID: 39708795
-
Activation of immune defences against parasitoid wasps does not underlie the cost of infection.Front Immunol. 2023 Dec 7;14:1275923. doi: 10.3389/fimmu.2023.1275923. eCollection 2023. Front Immunol. 2023. PMID: 38130722 Free PMC article.
-
MdSVWC1, a new pattern recognition receptor triggers multiple defense mechanisms against invading bacteria in Musca domestica.BMC Biol. 2024 Oct 23;22(1):242. doi: 10.1186/s12915-024-02042-5. BMC Biol. 2024. PMID: 39443921 Free PMC article.
-
The evolution of constitutively active humoral immune defenses in Drosophila populations under high parasite pressure.PLoS Pathog. 2024 Jan 11;20(1):e1011729. doi: 10.1371/journal.ppat.1011729. eCollection 2024 Jan. PLoS Pathog. 2024. PMID: 38206983 Free PMC article.
References
-
- Chapman S. J., Hill A. V., Human genetic susceptibility to infectious disease. Nat. Rev. Genet. 13, 175–188 (2012). - PubMed
-
- Salvaudon L., Giraud T., Shykoff J. A., Genetic diversity in natural populations: A fundamental component of plant-microbe interactions. Curr. Opin. Plant Biol. 11, 135–143 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
