

Transcription-Replication Conflicts as a Source of Genome Instability
- PMID: 37552891
- PMCID: PMC10760935
- DOI: 10.1146/annurev-genet-080320-031523
Transcription-Replication Conflicts as a Source of Genome Instability
Abstract
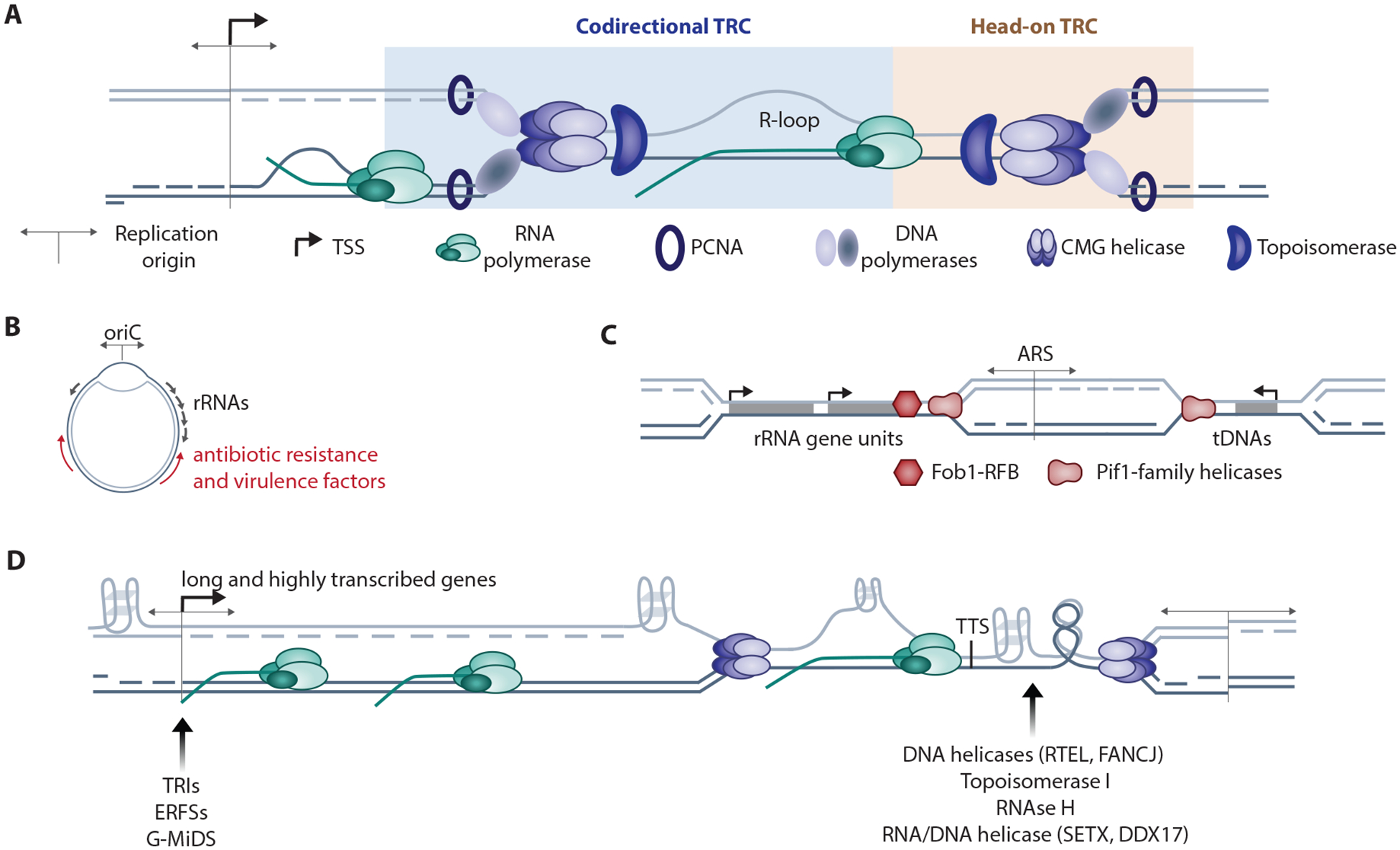
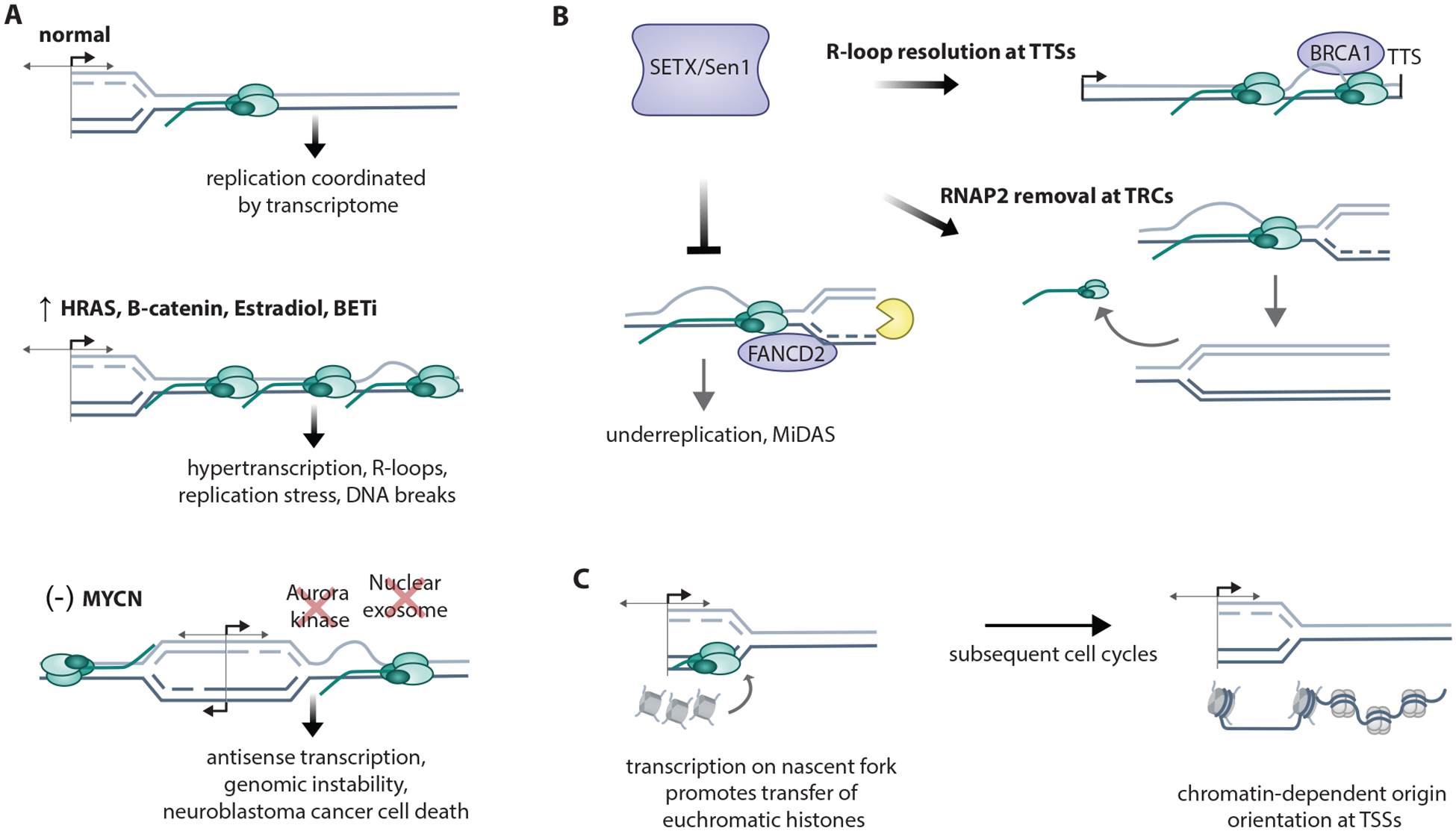
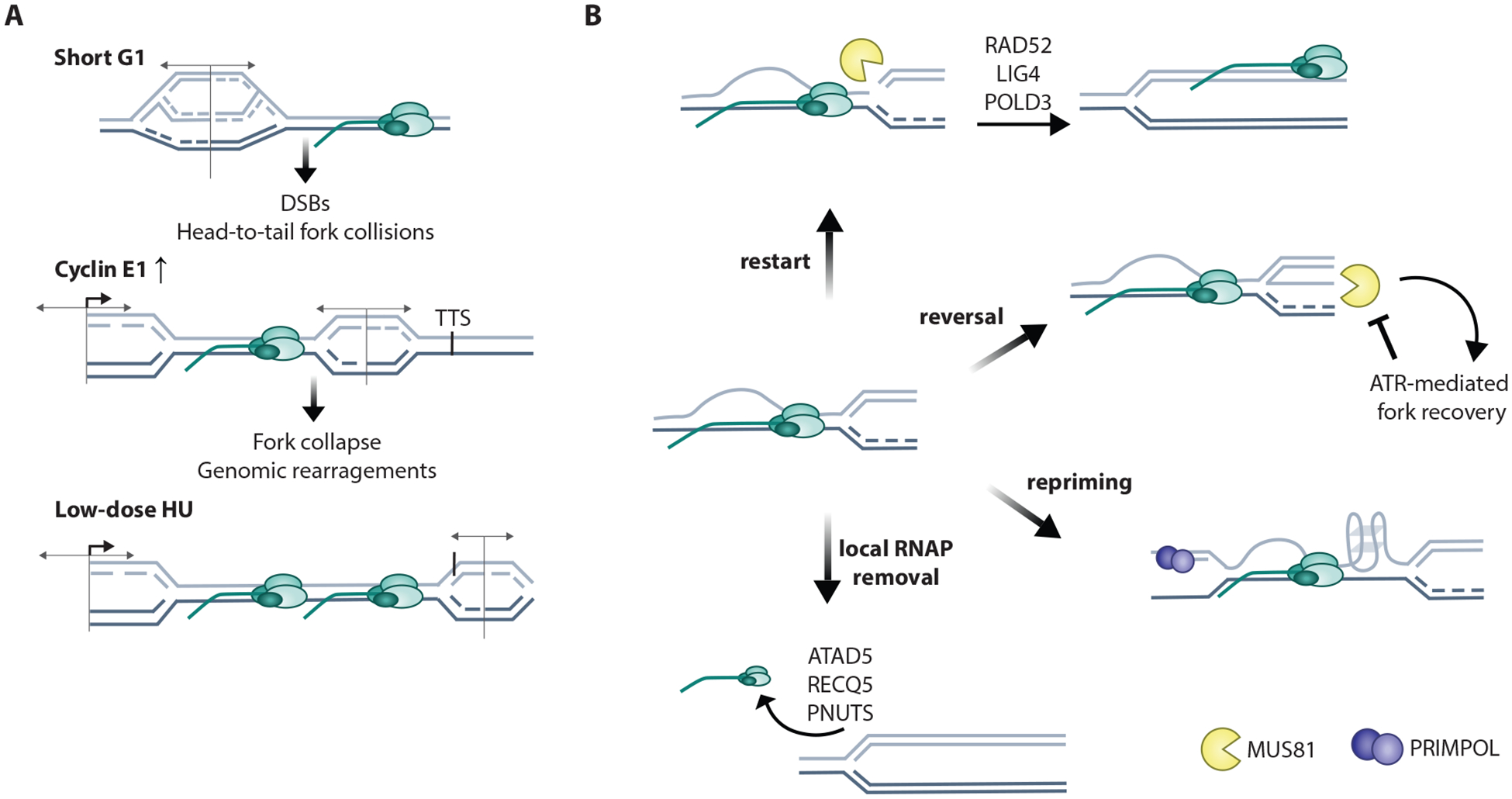
Transcription and replication both require large macromolecular complexes to act on a DNA template, yet these machineries cannot simultaneously act on the same DNA sequence. Conflicts between the replication and transcription machineries (transcription-replication conflicts, or TRCs) are widespread in both prokaryotes and eukaryotes and have the capacity to both cause DNA damage and compromise complete, faithful replication of the genome. This review will highlight recent studies investigating the genomic locations of TRCs and the mechanisms by which they may be prevented, mitigated, or resolved. We address work from both model organisms and mammalian systems but predominantly focus on multicellular eukaryotes owing to the additional complexities inherent in the coordination of replication and transcription in the context of cell type-specific gene expression and higher-order chromatin organization.
Keywords: DNA repair; RNA polymerase; TRCs; genomic instability; origin firing; replication stress; replisome; transcription; transcription–replication conflicts.
Figures
References
-
- Aiello U, Challal D, Wentzinger G, Lengronne A, Appanah R, et al. 2022. Sen1 is a key regulator of transcription-driven conflicts. Mol Cell. 82(16):2952–2966.e6 - PubMed
-
- Alvarez V, Bandau S, Jiang H, Rios-Szwed D, Hukelmann J, et al. 2023. Proteomic profiling reveals distinct phases to the restoration of chromatin following DNA replication. Cell Reports. 42(1):111996. - PubMed
-
- Bayona-Feliu A, Barroso S, Muñoz S, Aguilera A. 2021. The SWI/SNF chromatin remodeling complex helps resolve R-loop-mediated transcription–replication conflicts. Nat Genet, pp. 1–14 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
