

The mammalian midbody and midbody remnant are assembly sites for RNA and localized translation
- PMID: 37552987
- PMCID: PMC10592306
- DOI: 10.1016/j.devcel.2023.07.009
The mammalian midbody and midbody remnant are assembly sites for RNA and localized translation
Abstract
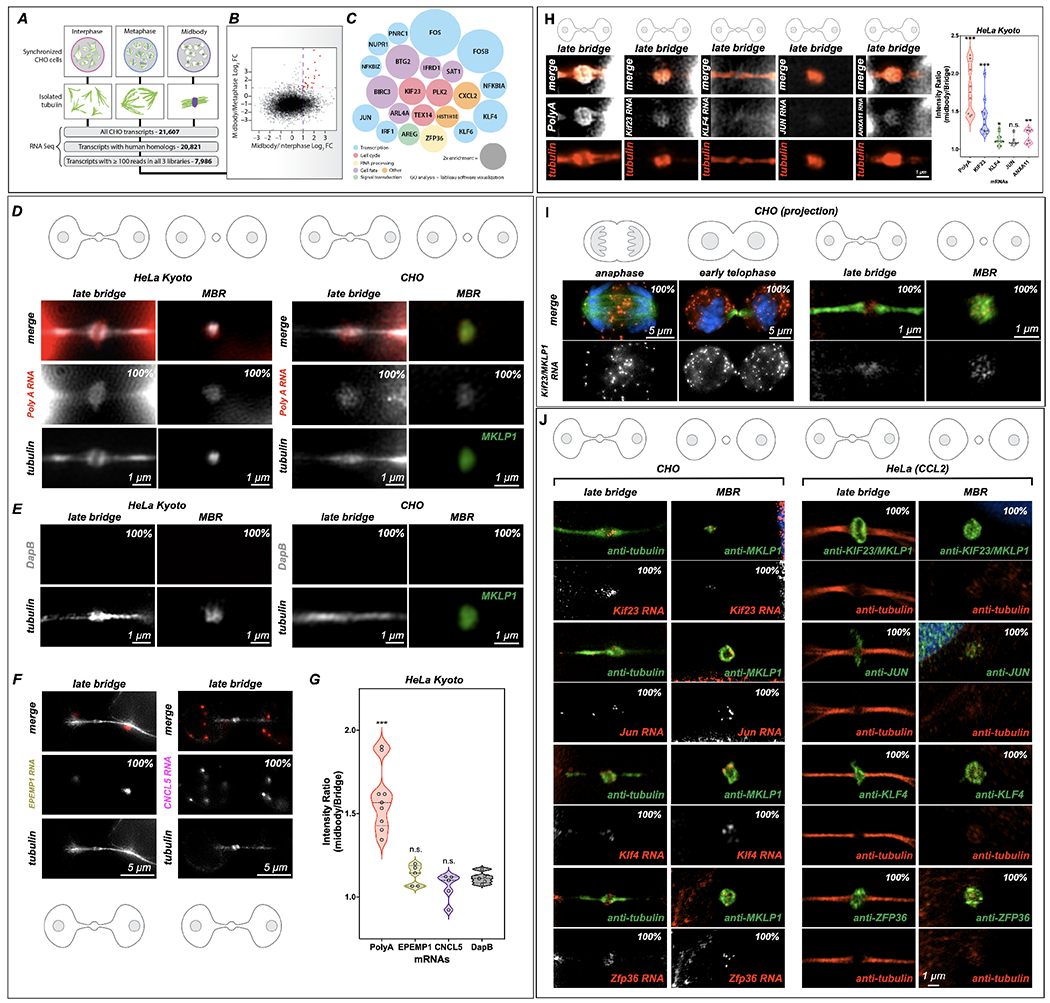
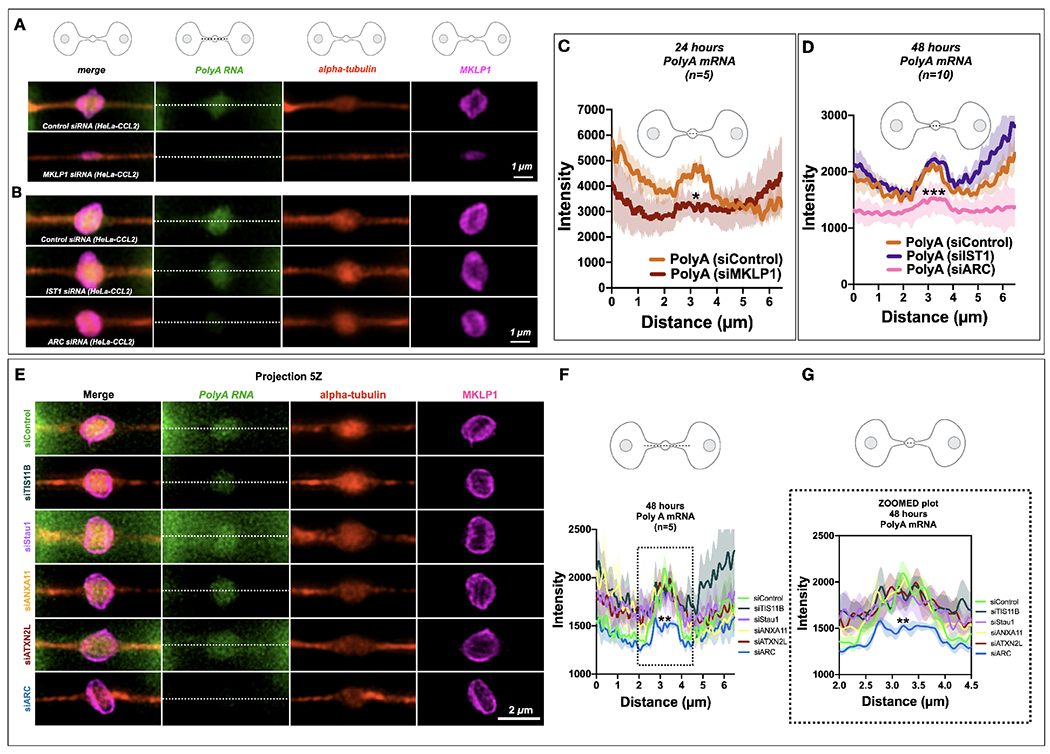
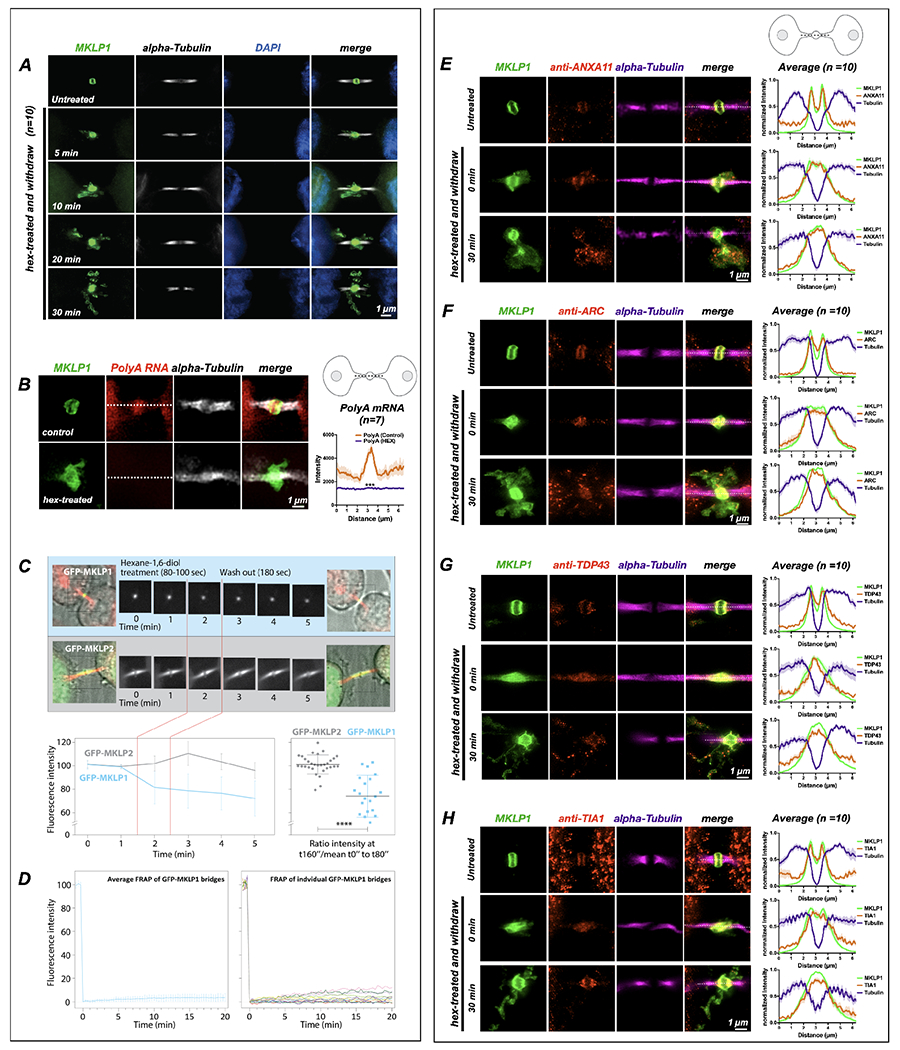
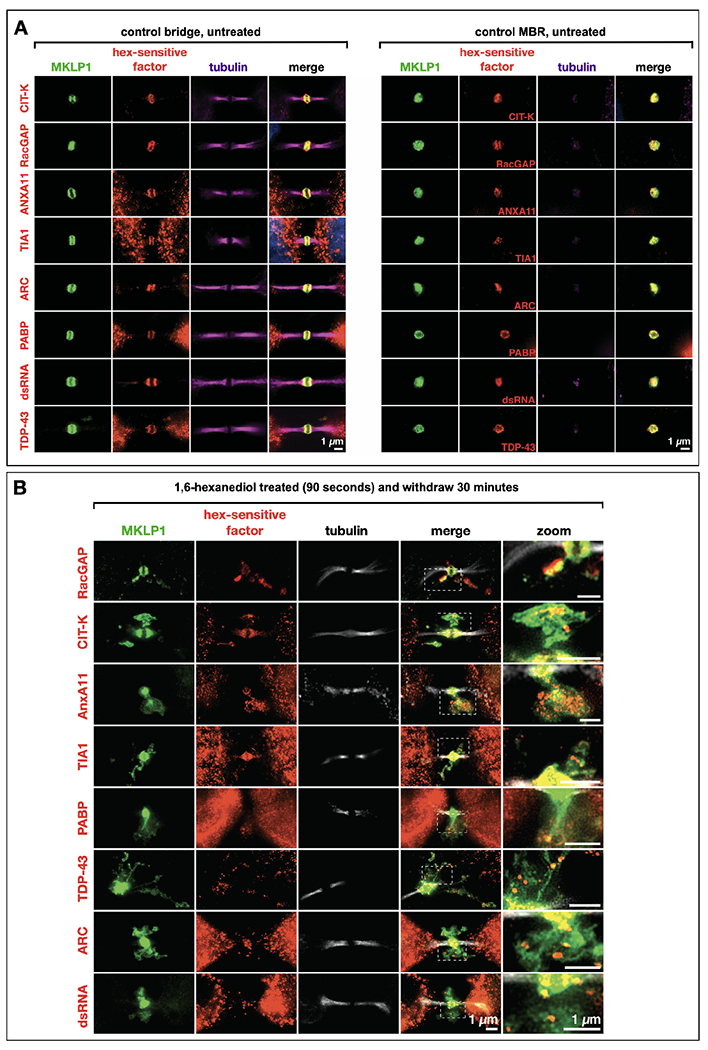
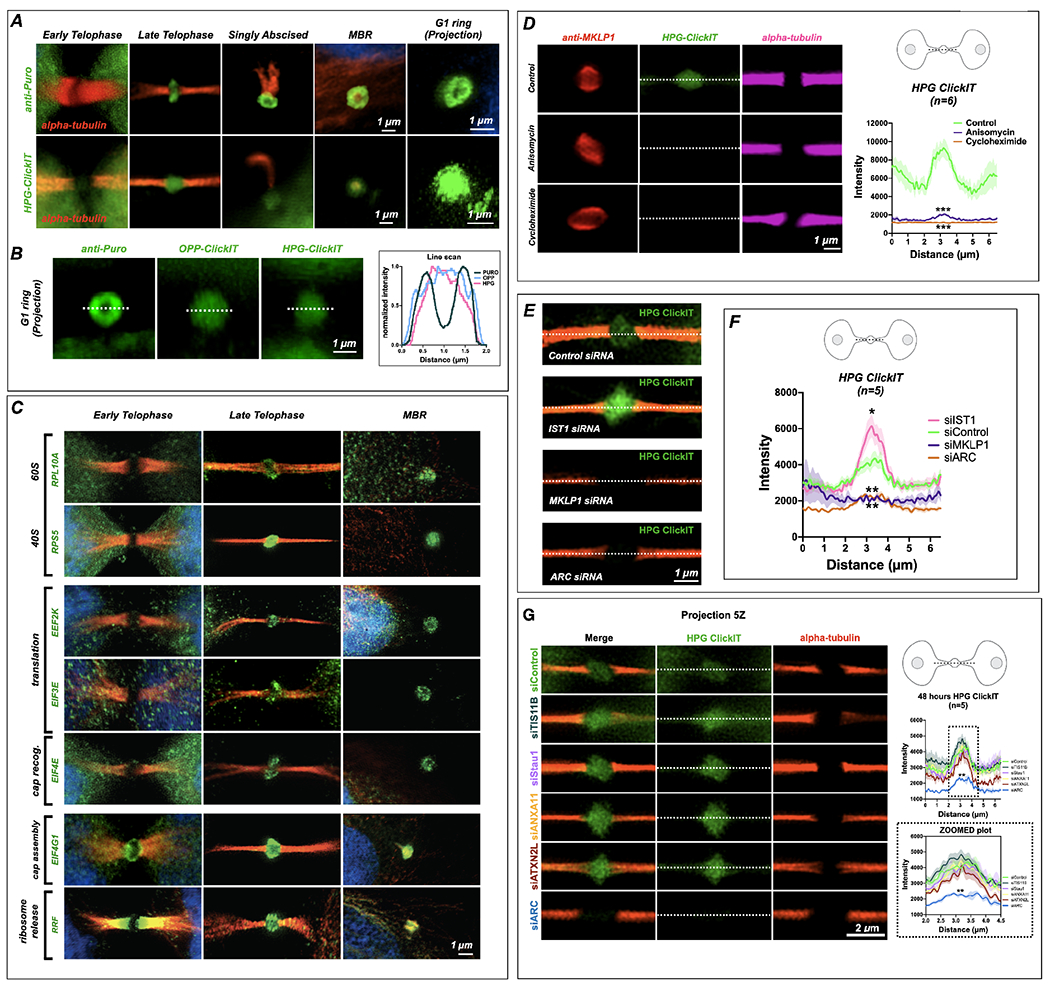
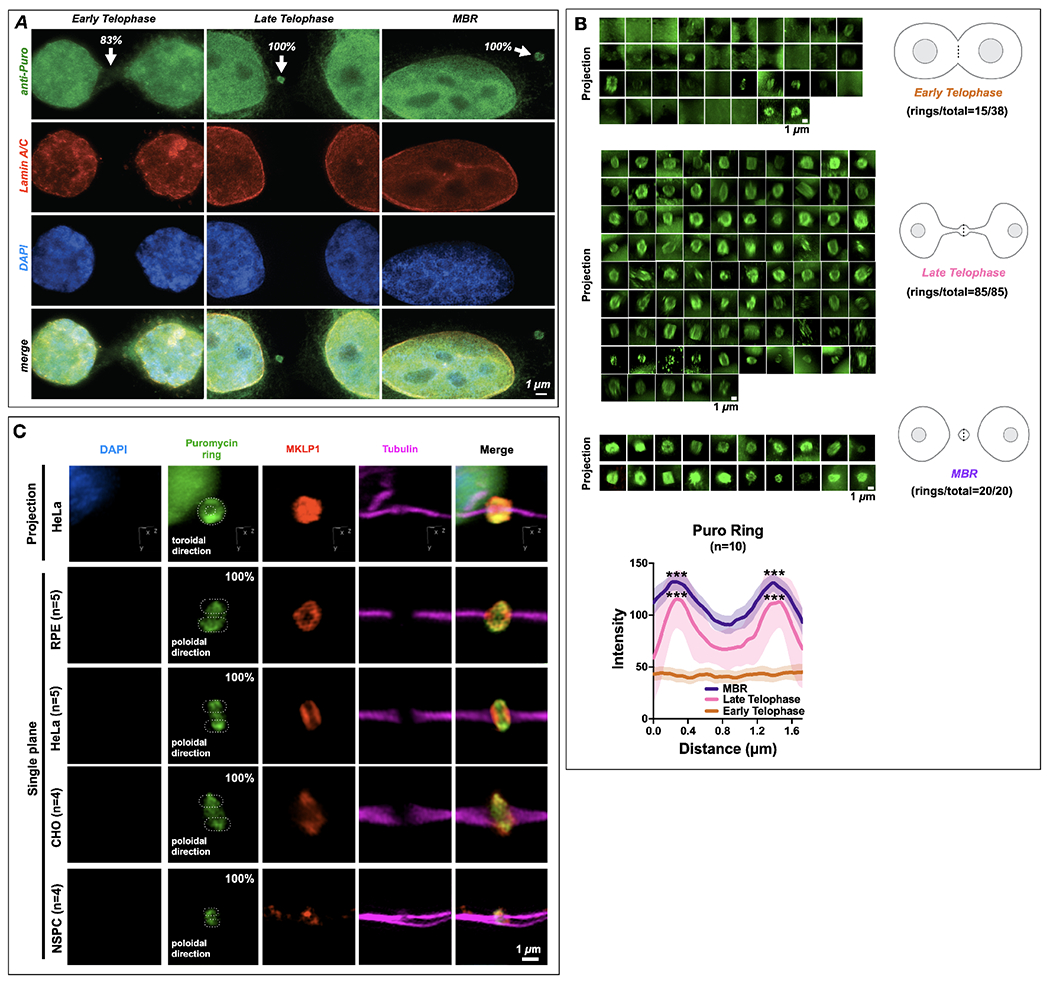
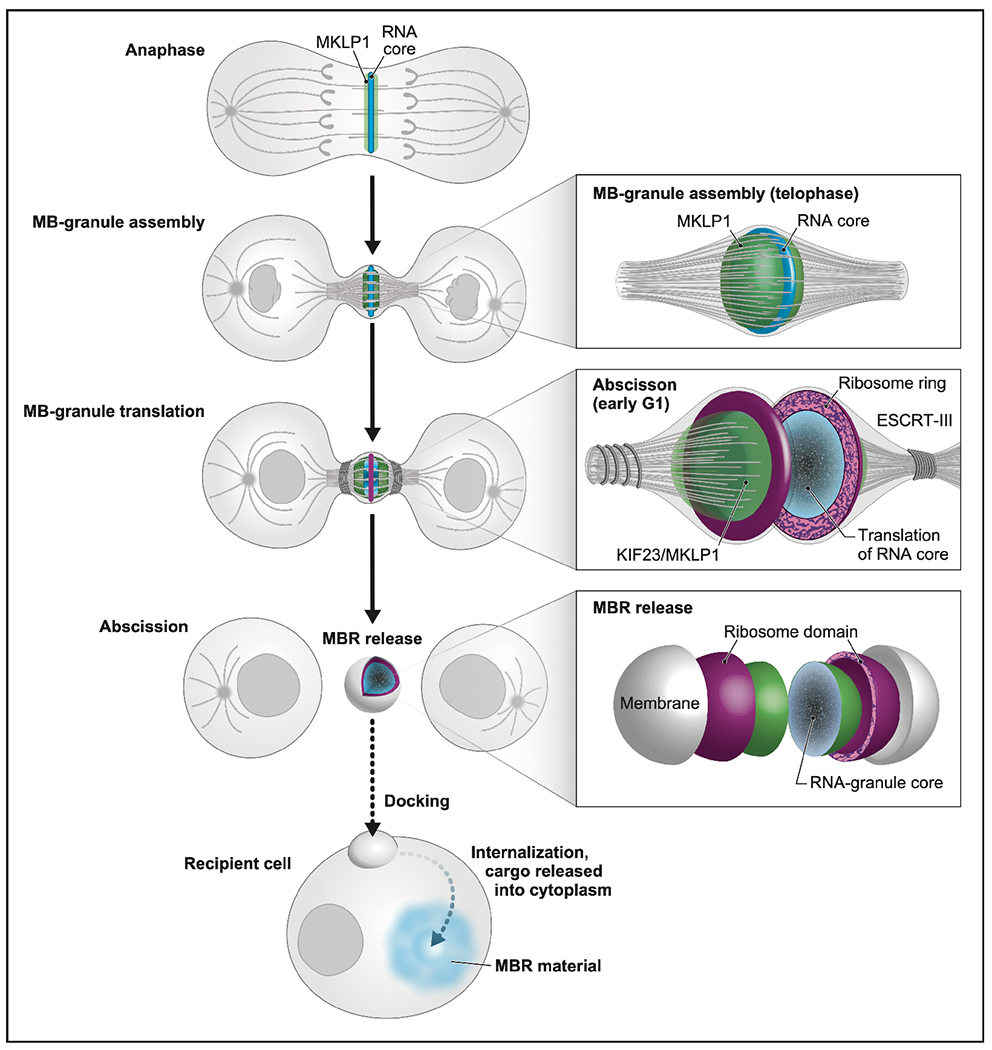
Long ignored as a vestigial remnant of cytokinesis, the mammalian midbody (MB) is released post-abscission inside large extracellular vesicles called MB remnants (MBRs). Recent evidence suggests that MBRs can modulate cell proliferation and cell fate decisions. Here, we demonstrate that the MB matrix is the site of ribonucleoprotein assembly and is enriched in mRNAs that encode proteins involved in cell fate, oncogenesis, and pluripotency, which we are calling the MB granule. Both MBs and post-abscission MBRs are sites of spatiotemporally regulated translation, which is initiated when nascent daughter cells re-enter G1 and continues after extracellular release. MKLP1 and ARC are necessary for the localization and translation of RNA in the MB dark zone, whereas ESCRT-III is necessary to maintain translation levels in the MB. Our work reveals a unique translation event that occurs during abscission and within a large extracellular vesicle.
Keywords: Arc; ESCRT-III; EV; MB; MBR; MBsome; RBP; abscission; cytokinesis; intercellular bridge; large extracellular vesicle; midbody; midbody remnant; mitosis; translation.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Mierzwa BE, Chiaruttini N, Redondo-Morata L, Filseck J.M. von, König J, Larios J, Poser I, Müller-Reichert T, Scheuring S, Roux A, et al. (2017). Dynamic subunit turnover in ESCRT-III assemblies is regulated by Vps4 to mediate membrane remodelling during cytokinesis. Nat Cell Biol 19, 787–798. 10.1038/ncb3559. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
