

Endothelial TAZ inhibits capillarization of liver sinusoidal endothelium and damage-induced liver fibrosis via nitric oxide production
- PMID: 37554269
- PMCID: PMC10405847
- DOI: 10.7150/thno.83714
Endothelial TAZ inhibits capillarization of liver sinusoidal endothelium and damage-induced liver fibrosis via nitric oxide production
Abstract
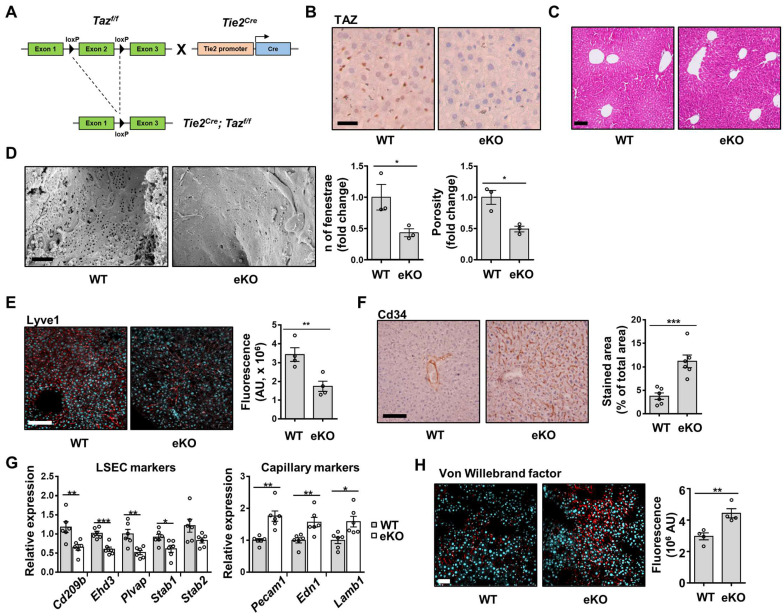
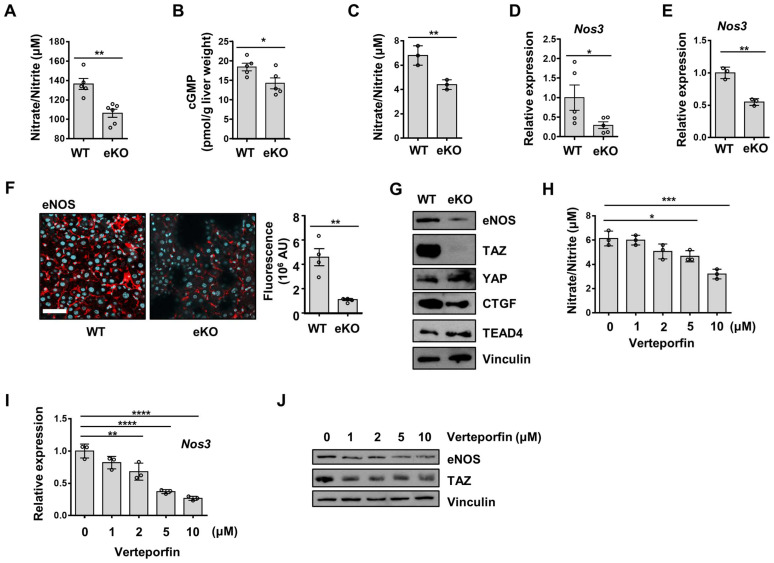
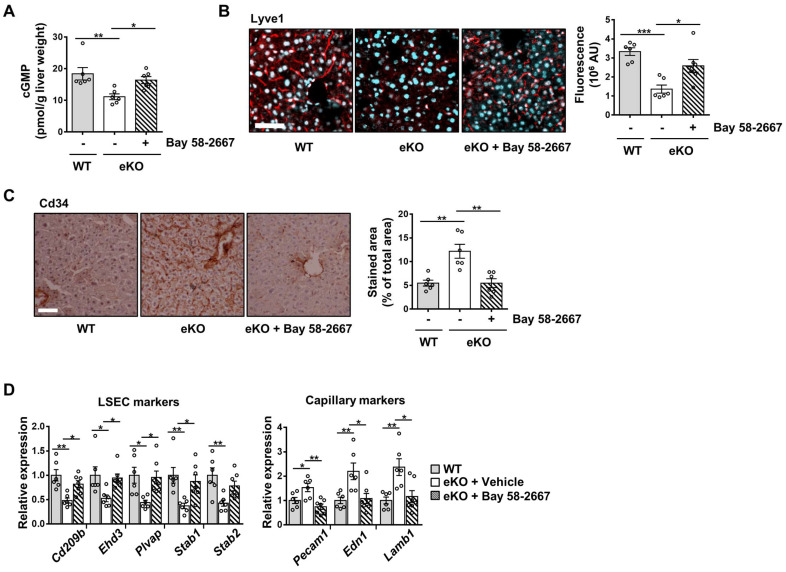
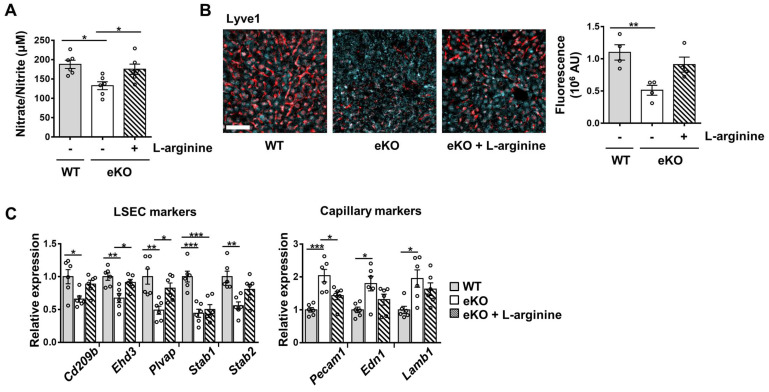
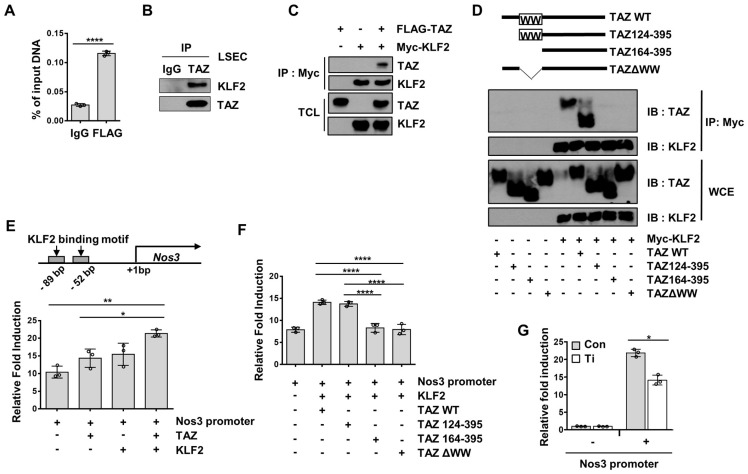
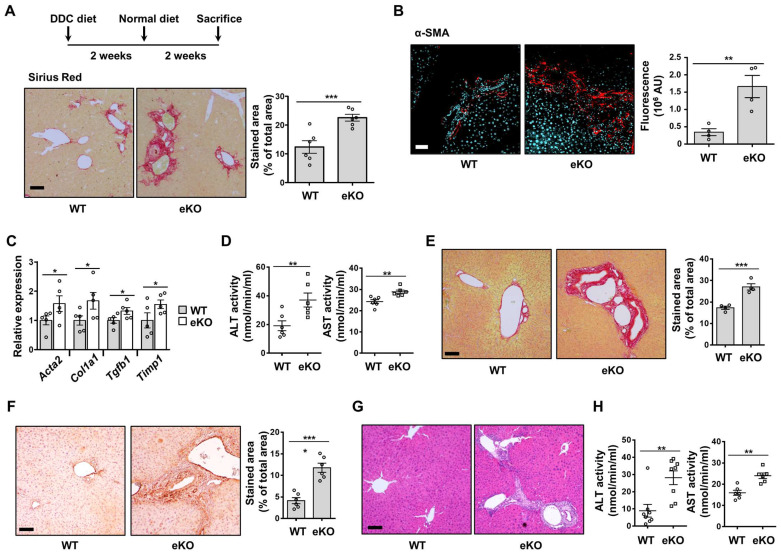
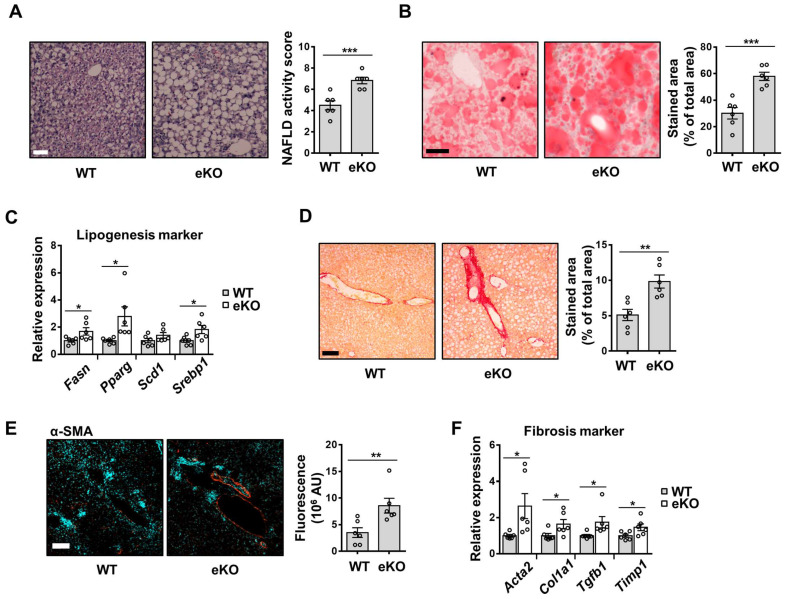
Background: Endothelial dysfunction is a systemic disorder and is involved in the pathogenesis of several human diseases. Hemodynamic shear stress plays an important role in vascular homeostasis including nitric oxide (NO) production. Impairment of NO production in endothelial cells stimulates the capillarization of liver sinusoidal endothelial cells, followed by hepatic stellate cell activation, inducing liver fibrosis. However, the detailed mechanism underlying NO production is not well understood. In hepatocytes, transcriptional co-activator with PDZ-binding motif (TAZ) has been reported to be involved in liver fibrosis. However, the role of endothelial TAZ in liver fibrosis has not been investigated. In this study, we uncovered the role TAZ in endothelial cell NO production, and its subsequent effects on liver fibrosis. Methods: TAZ-floxed mice were crossed with Tie2-cre transgenic mice, to generate endothelium-specific TAZ-knockout (eKO) mice. To induce liver damage, a 3,5-diethoxycarboncyl-1,4-dihydrocollidine, methionine-choline-deficient diet, or partial hepatectomy was applied. Liver fibrosis and endothelial dysfunction were analyzed in wild-type and eKO mice after liver damage. In addition, liver sinusoidal endothelial cell (LSEC) was used for in vitro assays of protein and mRNA levels. To study transcriptional regulation, chromatin immunoprecipitation and luciferase reporter assays were performed. Results: In liver of eKO mice, LSEC capillarization was observed, evidenced by loss of fenestrae and decreased LSEC-specific marker gene expression. LSEC capillarization of eKO mouse is caused by downregulation of endothelial nitric oxide synthase expression and subsequent decrease in NO concentration, which is transcriptionally regulated by TAZ-KLF2 binding to Nos3 promoter. Diminished NO concentration by TAZ knockout in endothelium accelerates liver fibrosis induced by liver damages. Conclusions: Endothelial TAZ inhibits damage-induced liver fibrosis via NO production. This highlights an unappreciated role of TAZ in vascular health and liver diseases.
Keywords: Endothelial dysfunction; Endothelial nitric oxide synthase; Liver fibrosis; Liver sinusoidal endothelial cells; Nitric oxide.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures
References
-
- Poisson J, Lemoinne S, Boulanger C, Durand F, Moreau R, Valla D. et al. Liver sinusoidal endothelial cells: Physiology and role in liver diseases. J Hepatol. 2017;66:212–27. - PubMed
-
- Gracia-Sancho J, Caparros E, Fernandez-Iglesias A, Frances R. Role of liver sinusoidal endothelial cells in liver diseases. Nature reviews Gastroenterology & hepatology. 2021;18:411–31. - PubMed
-
- Sorensen KK, Simon-Santamaria J, McCuskey RS, Smedsrod B. Liver Sinusoidal Endothelial Cells. Compr Physiol. 2015;5:1751–74. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
