

The deubiquitinase EIF3H promotes hepatocellular carcinoma progression by stabilizing OGT and inhibiting ferroptosis
- PMID: 37559097
- PMCID: PMC10413709
- DOI: 10.1186/s12964-023-01220-2
The deubiquitinase EIF3H promotes hepatocellular carcinoma progression by stabilizing OGT and inhibiting ferroptosis
Abstract
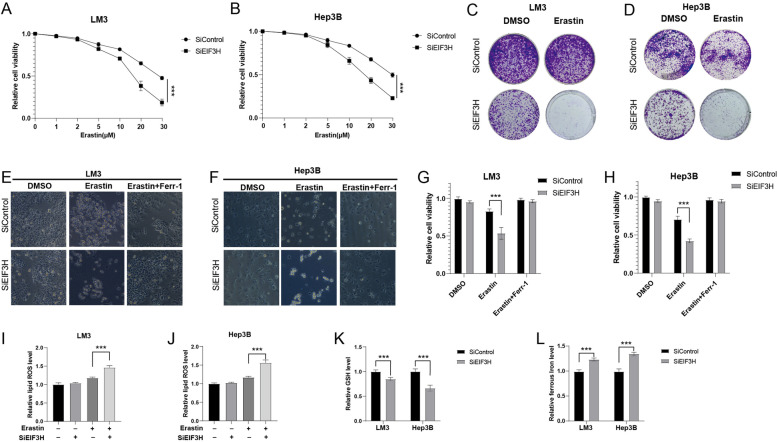
Hepatocellular carcinoma (HCC) is one of the most prevalent and lethal human malignancies, and with quite limited treatment alternatives. The proteasome is responsible for most of the protein degradation in eukaryotic cells and required for the maintenance of intracellular homeostasis. However, its potential role in HCC is largely unknown. In the current study, we identified eukaryotic translation initiation factor 3 subunit H (EIF3H), belonging to the JAB1/MPN/MOV34 (JAMM) superfamily, as a bona fide deubiquitylase of O-GlcNAc transferase (OGT) in HCC. We explored that EIF3H was positively associated with OGT in HCC and was related to the unfavorable prognosis. EIF3H could interact with, deubiquitylate, and stabilize OGT in a deubiquitylase-dependent manner. Specifically, EIF3H was associated with the GT domain of ERα via its JAB/MP domain, thus inhibiting the K48-linked ubiquitin chain on OGT. Besides, we demonstrated that the knockdown of EIF3H significantly reduced OGT protein expression, cell proliferation and invasion, and caused G1/S arrest of HCC. We also found that the deletion of EIF3H prompted ferroptosis in HCC cells. Finally, the effects of EIF3H depletion could be reversed by further OGT overexpression, implying that the OGT status is indispensable for EIF3H function in HCC carcinogenesis. In summary, our study described the oncogenic function of EIF3H and revealed an interesting post-translational mechanism between EIF3H, OGT, and ferroptosis in HCC. Targeting the EIF3H may be a promising approach in HCC. Video Abstract.
Keywords: Deubiquitination; EIF3H; Ferroptosis; Hepatocellular carcinoma; OGT.
© 2023. BioMed Central Ltd., part of Springer Nature.
Conflict of interest statement
The authors declare no competing interests.
Figures





Similar articles
-
Elevated expression of eukaryotic translation initiation factor 3H is associated with proliferation, invasion and tumorigenicity in human hepatocellular carcinoma.Oncotarget. 2016 Aug 2;7(31):49888-49901. doi: 10.18632/oncotarget.10222. Oncotarget. 2016. PMID: 27340783 Free PMC article.
-
N6-methyladenosine-mediated EIF3H promotes anaplastic thyroid cancer progression and ferroptosis resistance by stabilizing β-catenin.Free Radic Biol Med. 2025 Apr;231:38-47. doi: 10.1016/j.freeradbiomed.2025.02.025. Epub 2025 Feb 19. Free Radic Biol Med. 2025. PMID: 39984062
-
Targeting USP8 Inhibits O-GlcNAcylation of SLC7A11 to Promote Ferroptosis of Hepatocellular Carcinoma via Stabilization of OGT.Adv Sci (Weinh). 2023 Nov;10(33):e2302953. doi: 10.1002/advs.202302953. Epub 2023 Oct 22. Adv Sci (Weinh). 2023. PMID: 37867237 Free PMC article.
-
The deubiquitinating enzyme ATXN3 promotes hepatocellular carcinoma progression by stabilizing TAZ.Cancer Gene Ther. 2025 Jan;32(1):136-145. doi: 10.1038/s41417-024-00869-2. Epub 2024 Dec 13. Cancer Gene Ther. 2025. PMID: 39672915
-
Ferroptosis in hepatocellular carcinoma, from mechanism to effect.Front Oncol. 2024 Mar 5;14:1350011. doi: 10.3389/fonc.2024.1350011. eCollection 2024. Front Oncol. 2024. PMID: 38511140 Free PMC article. Review.
Cited by
-
O‑GlcNAcylation as an emerging molecular target for cholangiocarcinoma therapy (Review).Oncol Rep. 2025 Oct;54(4):119. doi: 10.3892/or.2025.8952. Epub 2025 Jul 19. Oncol Rep. 2025. PMID: 40682838 Free PMC article. Review.
-
Pharmacological Dissection Identifies Retatrutide Overcomes the Therapeutic Barrier of Obese TNBC Treatments through Suppressing the Interplay between Glycosylation and Ubiquitylation of YAP.Adv Sci (Weinh). 2025 Mar;12(11):e2407494. doi: 10.1002/advs.202407494. Epub 2025 Jan 27. Adv Sci (Weinh). 2025. PMID: 39868848 Free PMC article.
-
ULK1-dependent phosphorylation of OGT instructs the tumorigenicity of O-GlcNAcylation.Sci China Life Sci. 2025 Jun 25. doi: 10.1007/s11427-024-2924-6. Online ahead of print. Sci China Life Sci. 2025. PMID: 40571812
-
Targeting RPLP2 Triggers DLBCL Ferroptosis by Decreasing FXN Expression.Biomedicines. 2025 May 28;13(6):1320. doi: 10.3390/biomedicines13061320. Biomedicines. 2025. PMID: 40564039 Free PMC article.
-
KDM2B regulates stroke injury by modulating OGT-mediated 0-GlcNAcylation of SLC7A11.Commun Biol. 2024 Nov 18;7(1):1530. doi: 10.1038/s42003-024-07251-w. Commun Biol. 2024. PMID: 39558086 Free PMC article.
References
-
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA: a cancer journal for clinicians 2022;72:7–33. 10.3322/caac.21708. - PubMed
-
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of incidence and mortality worldwide for 36 Cancers in 185 Countries. CA: a cancer j clinic 2021;71:209–249. 10.3322/caac.21660. - PubMed
-
- Heimbach, J.K.; Kulik, L.M.; Finn, R.S.; Sirlin, C.B.; Abecassis, M.M.; Roberts, L.R.; Zhu, A.X.; Murad, M.H.; Marrero, J.A. AASLD guidelines for the treatment of hepatocellular carcinoma. Hepatology (Baltimore, Md.) 2018;67:358–380. 10.1002/hep.29086. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
