

Past energy allocation overwhelms current energy stresses in determining energy allocation trade-offs
- PMID: 37560183
- PMCID: PMC10408252
- DOI: 10.1002/ece3.10402
Past energy allocation overwhelms current energy stresses in determining energy allocation trade-offs
Abstract
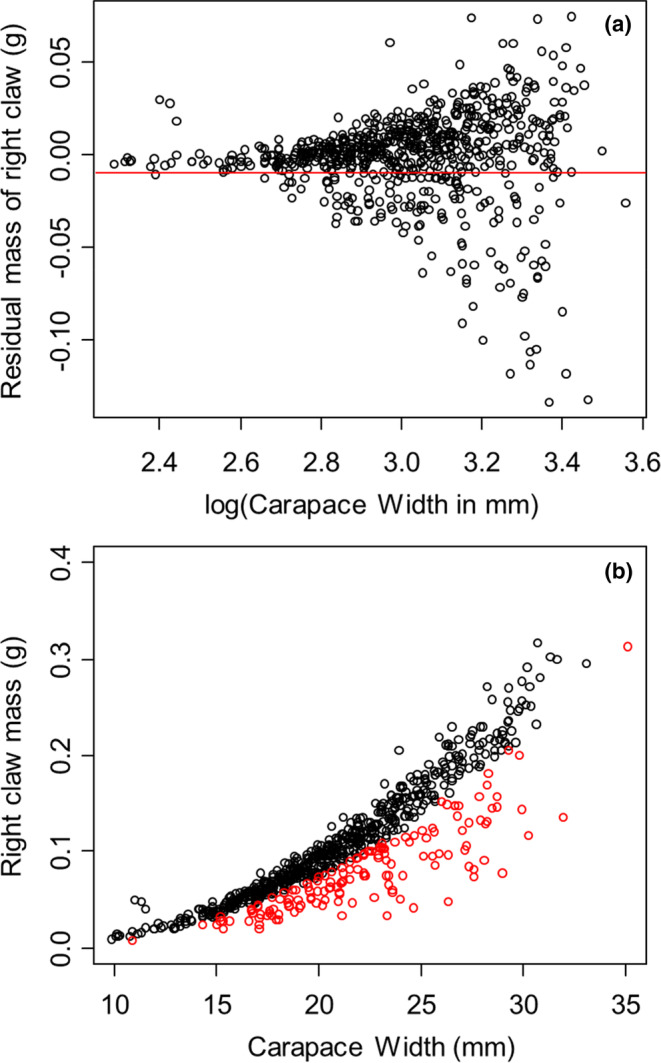
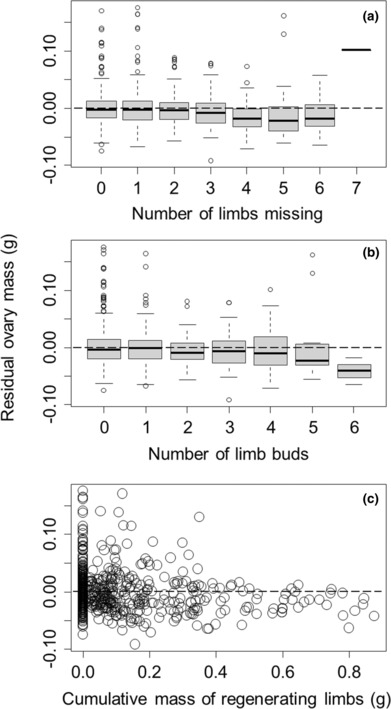
Regeneration of lost appendages is a gradual process in many species, spreading energetic costs of regeneration through time. Energy allocated to the regeneration of lost appendages cannot be used for other purposes and, therefore, commonly elicits energetic trade-offs in biological processes. We used limb loss in the Asian shore crab Hemigrapsus sanguineus to compare the strength of energetic trade-offs resulting from historic limb losses that have been partially regenerated versus current injuries that have not yet been repaired. Consistent with previous studies, we show that limb loss and regeneration results in trade-offs that reduce reproduction, energy storage, and growth. As may be expected, we show that trade-offs in these metrics from historic limb losses far outweigh trade-offs from current limb losses, and correlate directly with the degree of historic limb loss that has been regenerated. As regenerating limbs get closer to their normal size, these historical injuries get harder to detect, despite the continued allocation of additional resources to limb development. Our results demonstrate the importance of and a method for identifying historic appendage losses and of quantifying the amount of regeneration that has already occurred, as opposed to assessing only current injury, to accurately assess the strength of energetic trade-offs in animals recovering from nonlethal injury.
Keywords: Asian shore crabs; Hemigrapsus sanguineus; energetic trade‐offs; limb regeneration; nonlethal injury; regeneration.
© 2023 The Authors. Ecology and Evolution published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures



References
-
- Baranowitz, S. A. , Maderson, P. F. A. , & Connelly, T. G. (1979). Lizard and newt tail regeneration: A quantitative study. Journal of Experimental Zoology, 210(1), 17–37. - PubMed
-
- Bateman, P. W. , & Fleming, P. A. (2009). To cut a long tail short: A review of lizard caudal autotomy studies carried out over the last 20 years. Journal of Zoology, 277(1), 1–14.
-
- Bely, A. E. (2010). Evolutionary loss of animal regeneration: Pattern and process. Integrative and Comparative Biology, 50(4), 515–527. - PubMed
-
- Bingham, B. L. , Burr, J. , & Head, H. W. (2000). Causes and consequences of arm damage in the sea star Leptasterias hexactis . Canadian Journal of Zoology, 78(4), 596–605.
-
- Buechley, E. R. , Oppel, S. , Efrat, R. , Phipps, W. L. , Carbonell Alanis, I. , Álvarez, E. , Andreotti, A. , Arkumarev, V. , Berger‐Tal, O. , Bermejo Bermejo, A. , Bounas, A. , Ceccolini, G. , Cenerini, A. , Dobrev, V. , Duriez, O. , García, J. , García‐Ripollés, C. , Galán, M. , Gil, A. , … Marra, P. P. (2021). Differential survival throughout the full annual cycle of a migratory bird presents a life‐history trade‐off. Journal of Animal Ecology, 90(5), 1228–1238. - PubMed
Associated data
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
