

Replication dynamics identifies the folding principles of the inactive X chromosome
- PMID: 37563439
- PMCID: PMC10442229
- DOI: 10.1038/s41594-023-01052-1
Replication dynamics identifies the folding principles of the inactive X chromosome
Abstract
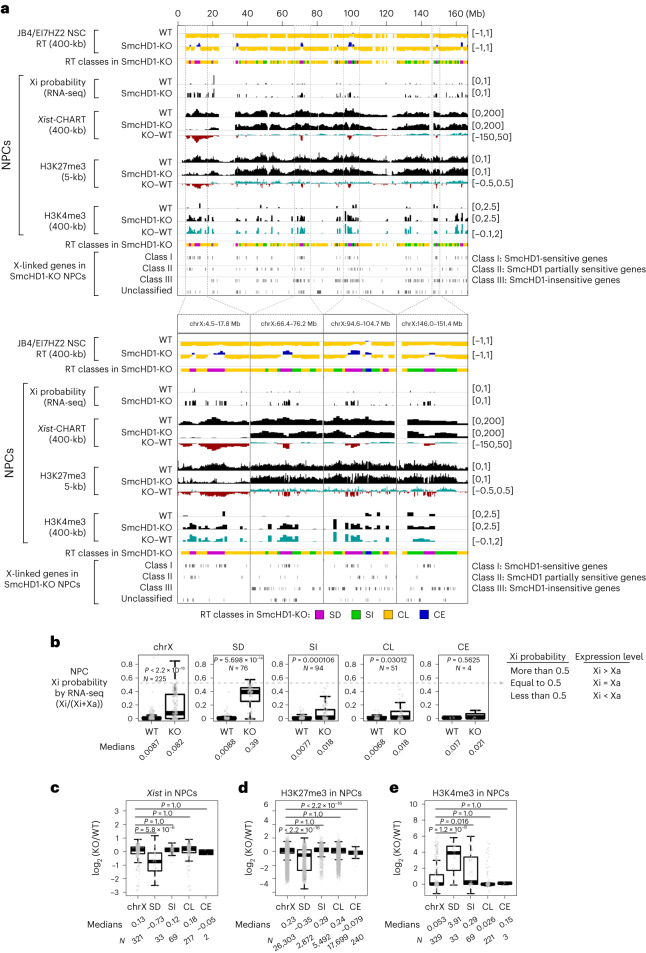
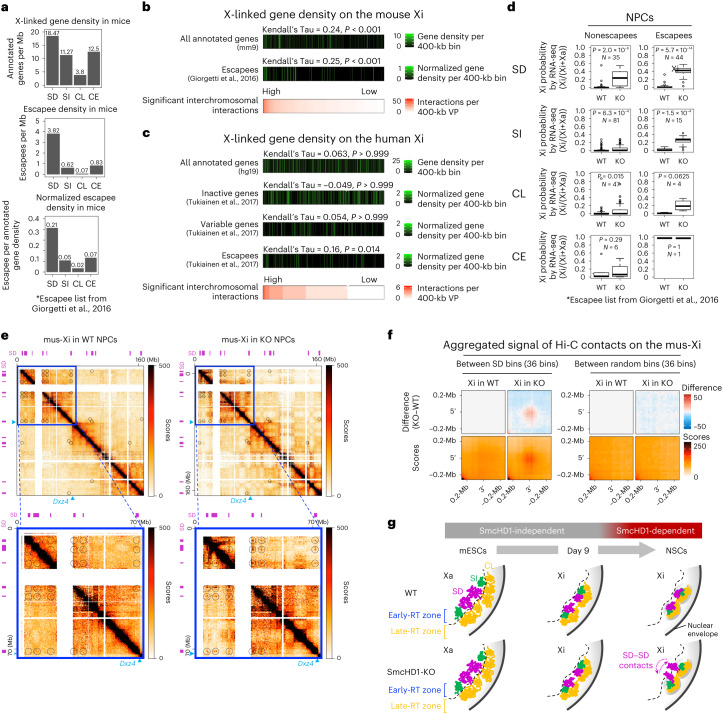
Chromosome-wide late replication is an enigmatic hallmark of the inactive X chromosome (Xi). How it is established and what it represents remains obscure. By single-cell DNA replication sequencing, here we show that the entire Xi is reorganized to replicate rapidly and uniformly in late S-phase during X-chromosome inactivation (XCI), reflecting its relatively uniform structure revealed by 4C-seq. Despite this uniformity, only a subset of the Xi became earlier replicating in SmcHD1-mutant cells. In the mutant, these domains protruded out of the Xi core, contacted each other and became transcriptionally reactivated. 4C-seq suggested that they constituted the outermost layer of the Xi even before XCI and were rich in escape genes. We propose that this default positioning forms the basis for their inherent heterochromatin instability in cells lacking the Xi-binding protein SmcHD1 or exhibiting XCI escape. These observations underscore the importance of 3D genome organization for heterochromatin stability and gene regulation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
