

An anchor-tether 'hindered' HCN1 inhibitor is antihyperalgesic in a rat spared nerve injury neuropathic pain model
- PMID: 37567808
- PMCID: PMC10541997
- DOI: 10.1016/j.bja.2023.06.067
An anchor-tether 'hindered' HCN1 inhibitor is antihyperalgesic in a rat spared nerve injury neuropathic pain model
Abstract
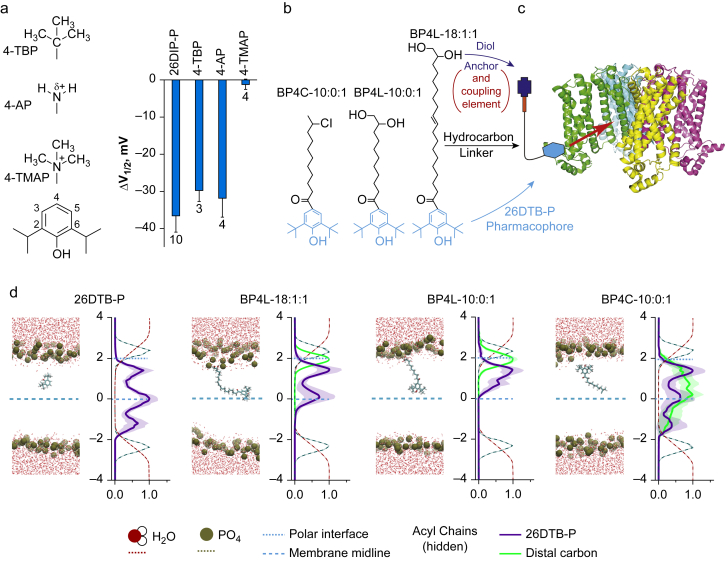
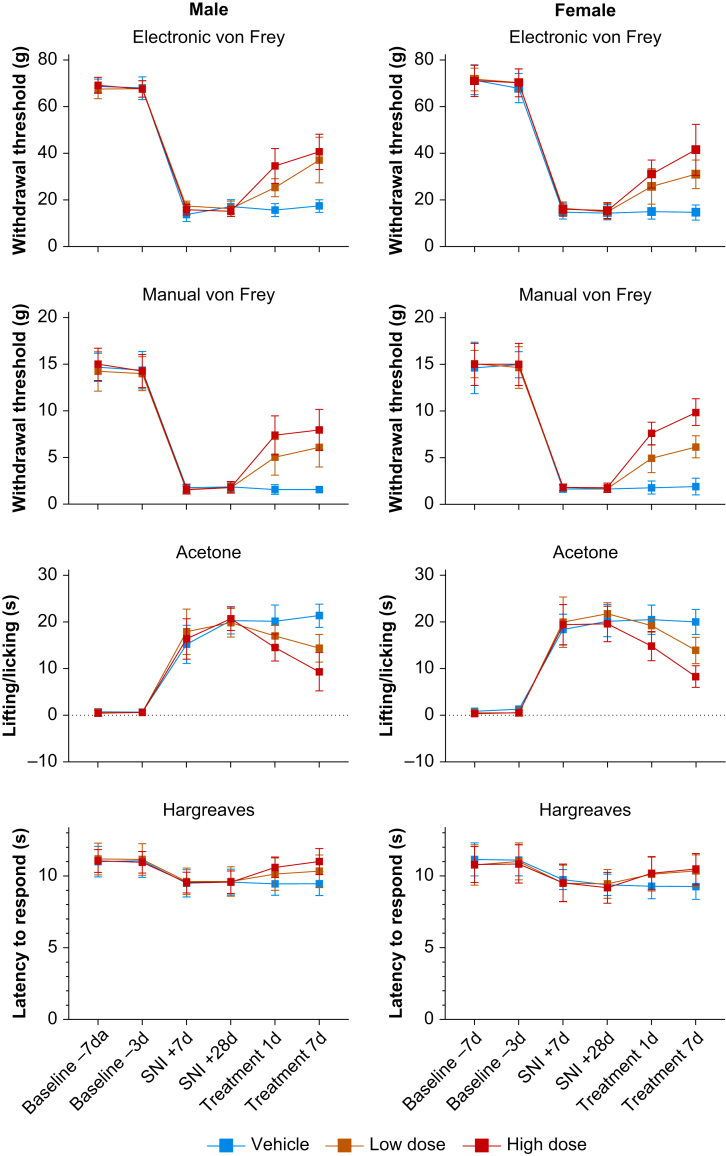
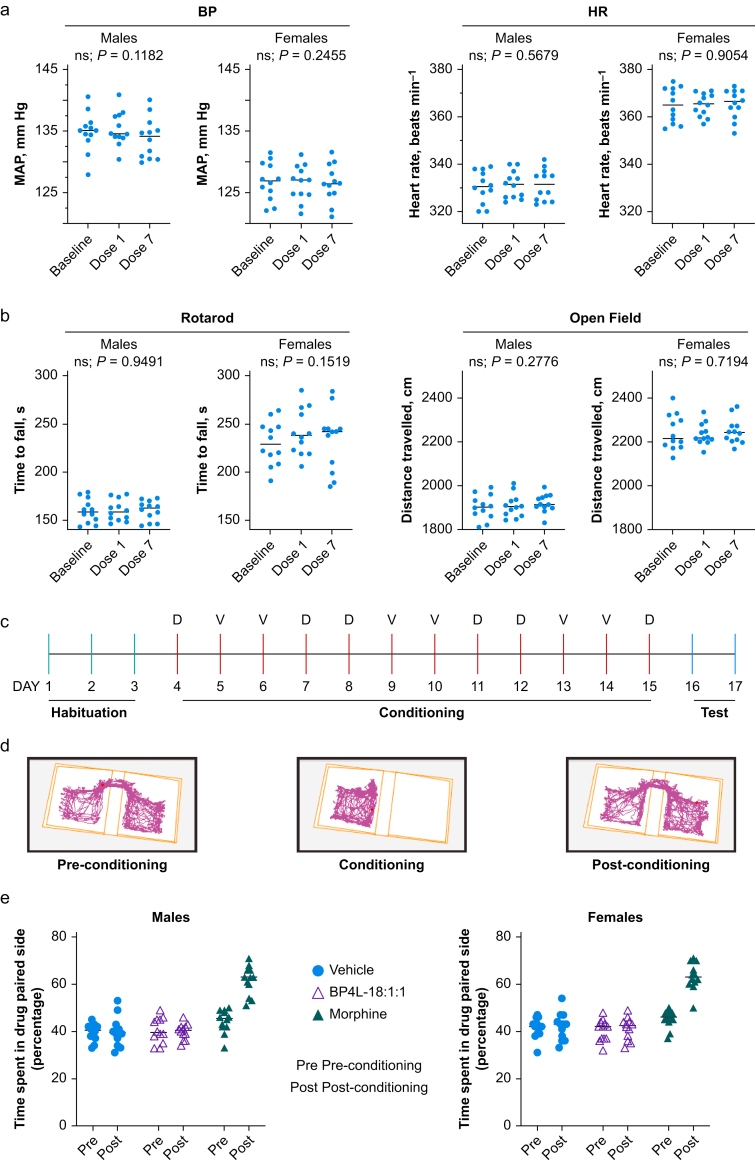
Background: Neuropathic pain impairs quality of life, is widely prevalent, and incurs significant costs. Current pharmacological therapies have poor/no efficacy and significant adverse effects; safe and effective alternatives are needed. Hyperpolarisation-activated cyclic nucleotide-regulated (HCN) channels are causally implicated in some forms of peripherally mediated neuropathic pain. Whilst 2,6-substituted phenols, such as 2,6-di-tert-butylphenol (26DTB-P), selectively inhibit HCN1 gating and are antihyperalgesic, the development of therapeutically tolerable, HCN-selective antihyperalgesics based on their inverse agonist activity requires that such drugs spare the cardiac isoforms and do not cross the blood-brain barrier.
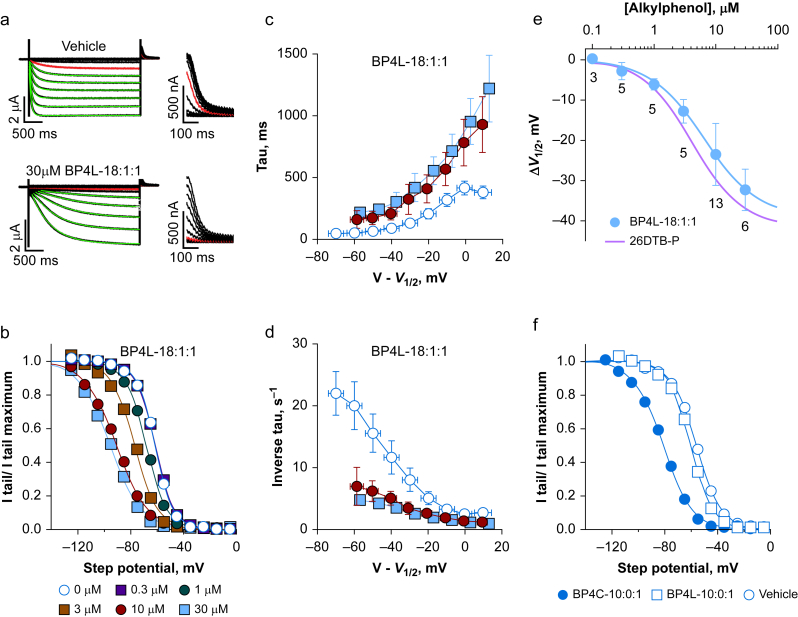
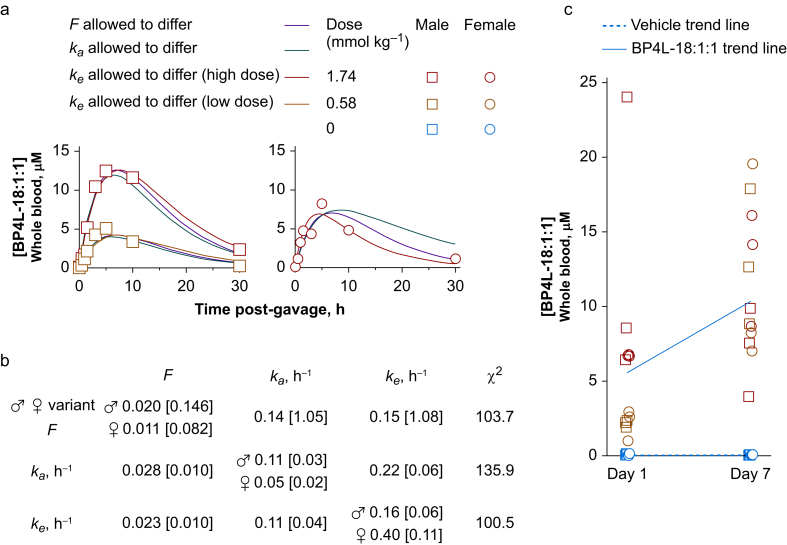
Methods: In silico molecular dynamics simulation, in vitro electrophysiology, and in vivo rat spared nerve injury methods were used to test whether 'hindered' variants of 26DTB-P (wherein a hydrophilic 'anchor' is attached in the para-position of 26DTB-P via an acyl chain 'tether') had the desired properties.
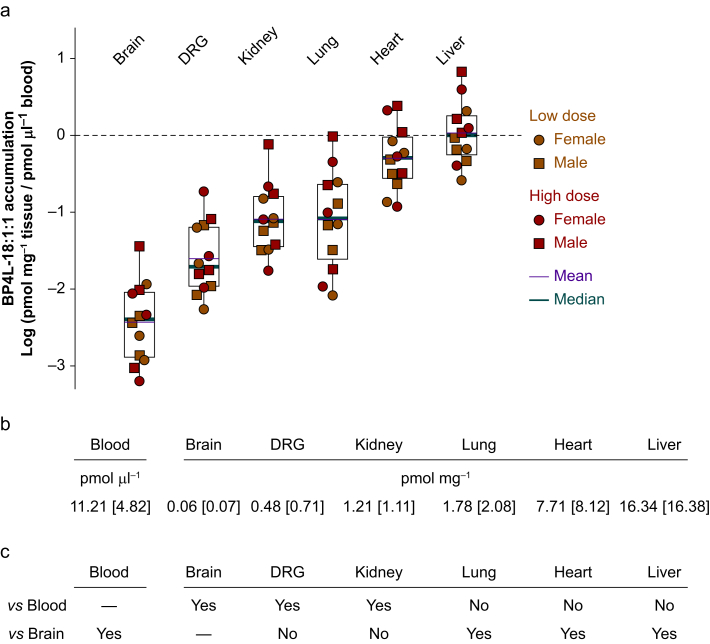
Results: Molecular dynamics simulation showed that membrane penetration of hindered 26DTB-Ps is controlled by a tethered diol anchor without elimination of head group rotational freedom. In vitro and in vivo analysis showed that BP4L-18:1:1, a variant wherein a diol anchor is attached to 26DTB-P via an 18-carbon tether, is an HCN1 inverse agonist and an orally available antihyperalgesic. With a CNS multiparameter optimisation score of 2.25, a >100-fold lower drug load in the brain vs blood, and an absence of adverse cardiovascular or CNS effects, BP4L-18:1:1 was shown to be poorly CNS penetrant and cardiac sparing.
Conclusions: These findings provide a proof-of-concept demonstration that anchor-tethered drugs are a new chemotype for treatment of disorders involving membrane targets.
Keywords: HCN1; antihyperalgesia; ion channel; nerve injury; neuropathic pain; rat.
Copyright © 2023 British Journal of Anaesthesia. Published by Elsevier Ltd. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
