

Regulation of DNA damage and transcriptional output in the vasculature through a cytoglobin-HMGB2 axis
- PMID: 37573836
- PMCID: PMC10428073
- DOI: 10.1016/j.redox.2023.102838
Regulation of DNA damage and transcriptional output in the vasculature through a cytoglobin-HMGB2 axis
Abstract
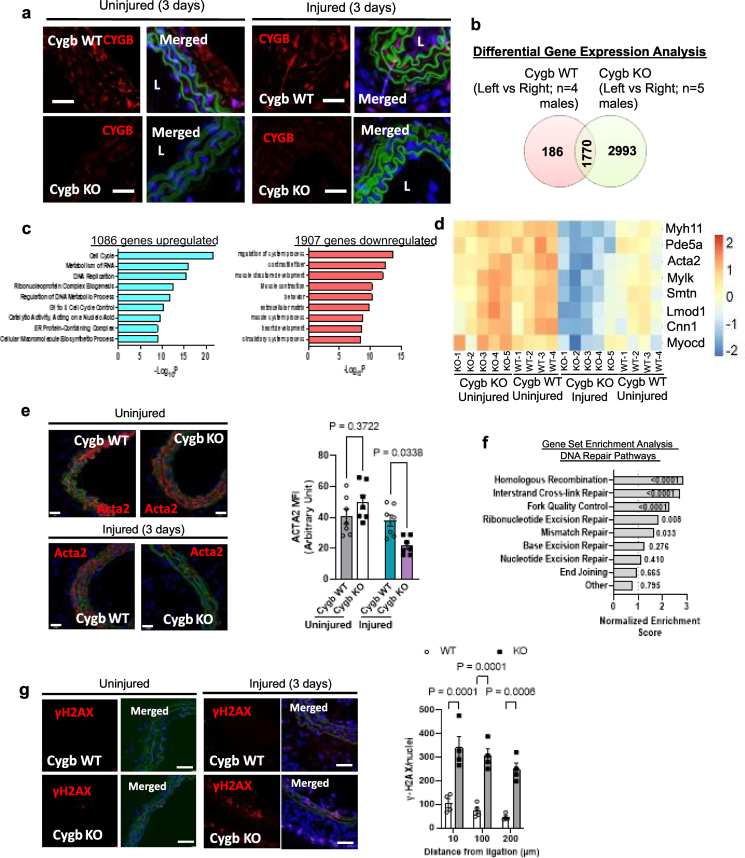
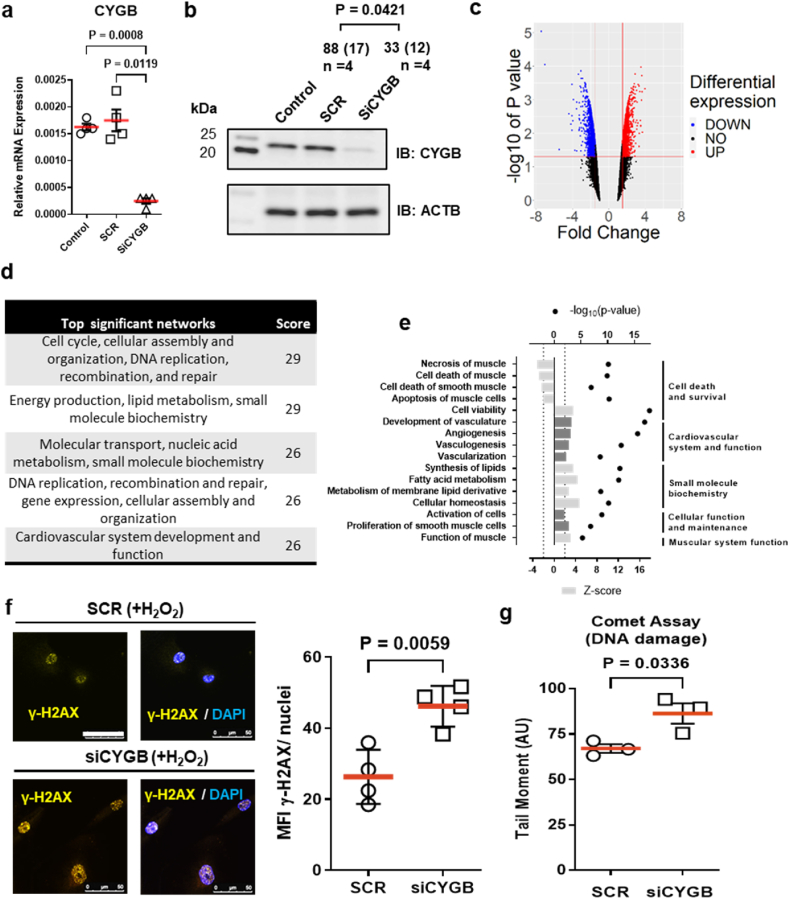
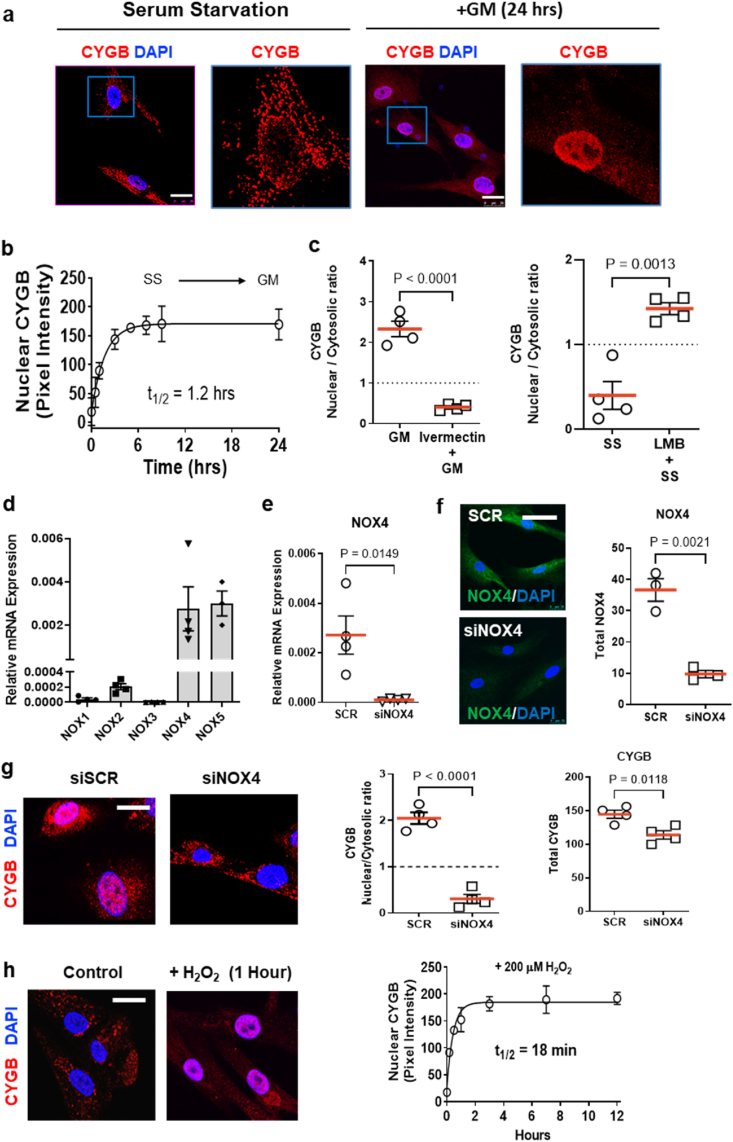
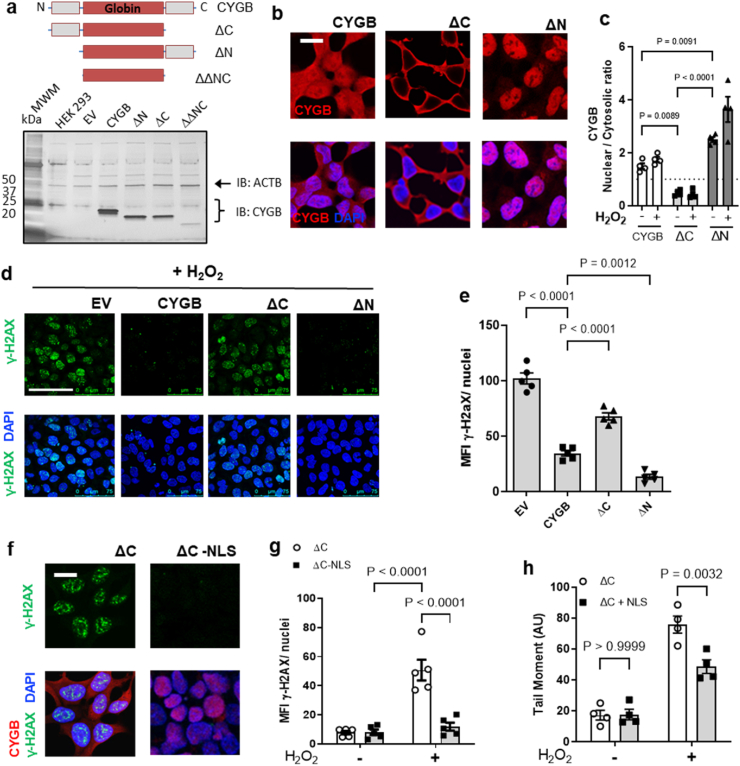
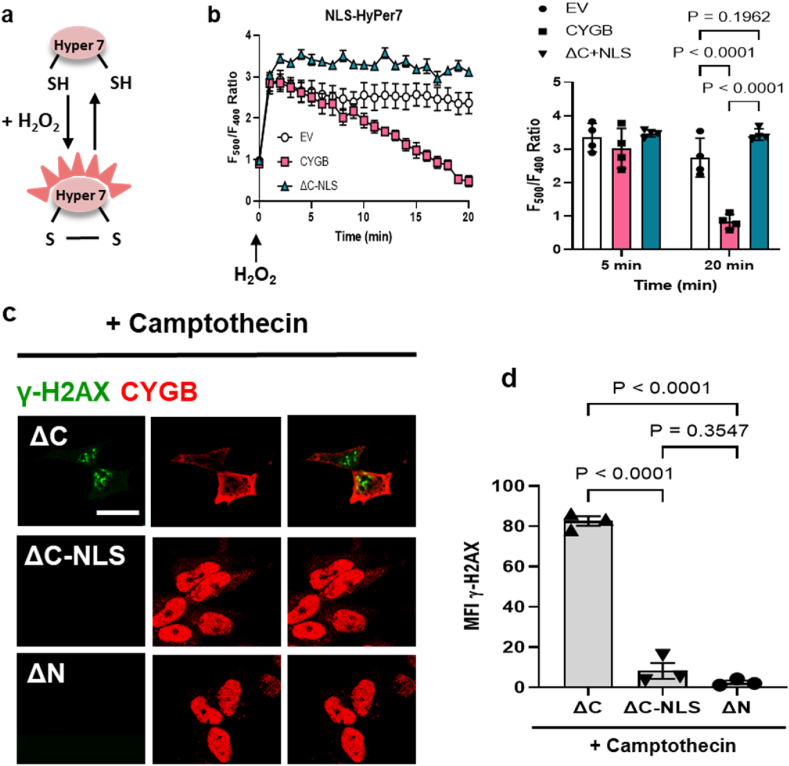
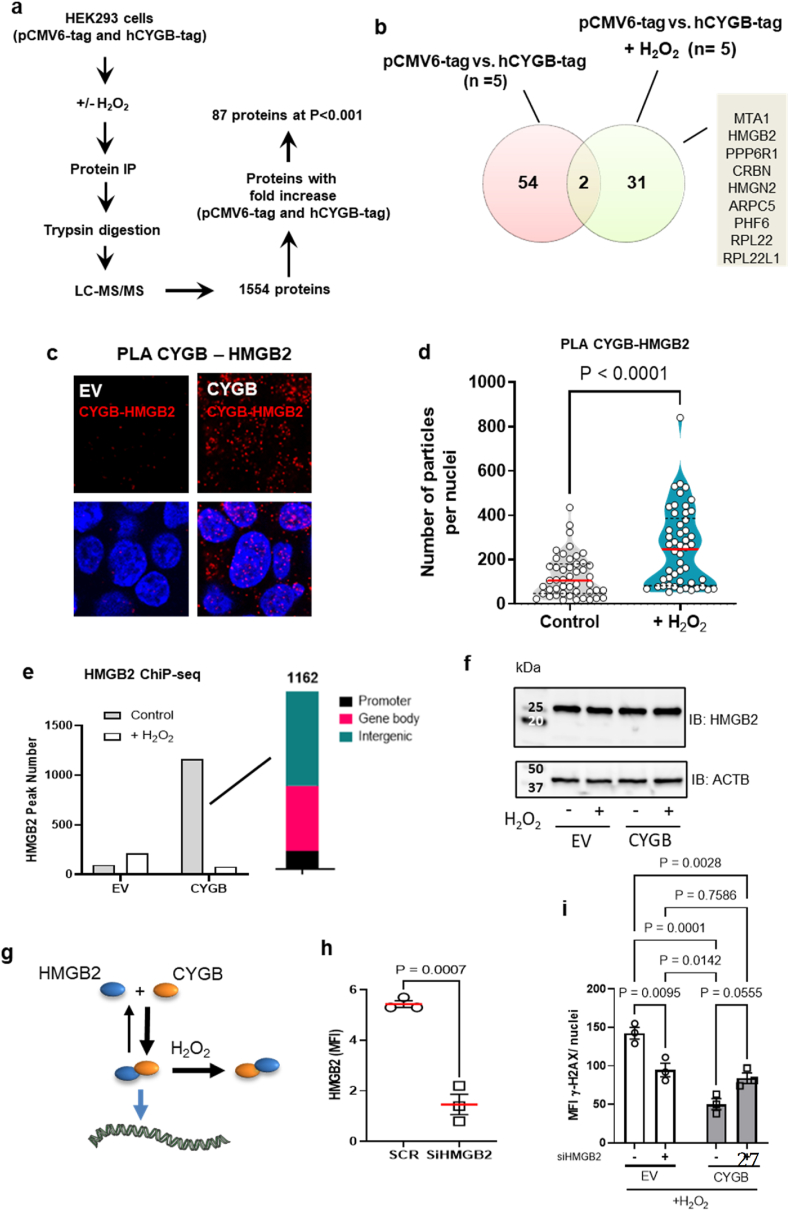
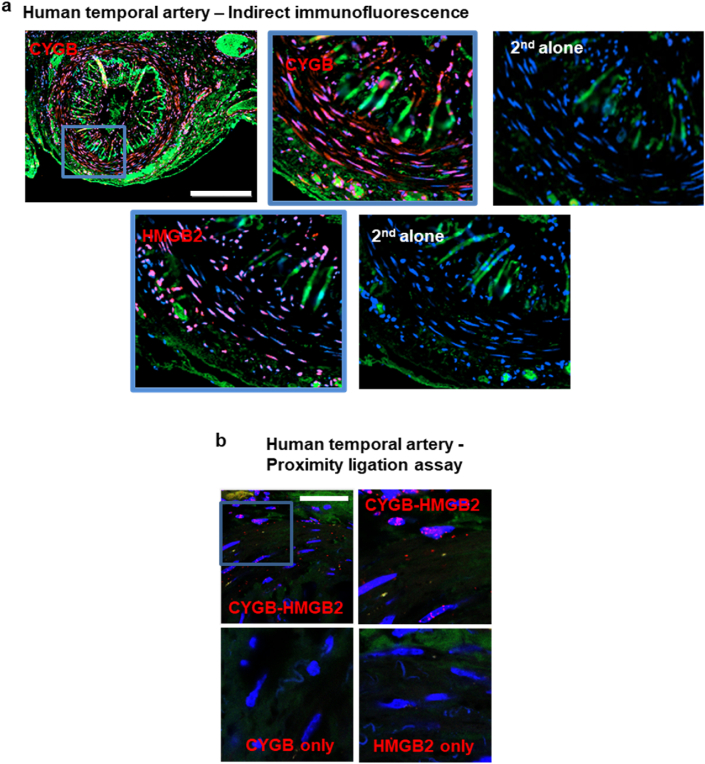
Identifying novel regulators of vascular smooth muscle cell function is necessary to further understand cardiovascular diseases. We previously identified cytoglobin, a hemoglobin homolog, with myogenic and cytoprotective roles in the vasculature. The specific mechanism of action of cytoglobin is unclear but does not seem to be related to oxygen transport or storage like hemoglobin. Herein, transcriptomic profiling of injured carotid arteries in cytoglobin global knockout mice revealed that cytoglobin deletion accelerated the loss of contractile genes and increased DNA damage. Overall, we show that cytoglobin is actively translocated into the nucleus of vascular smooth muscle cells through a redox signal driven by NOX4. We demonstrate that nuclear cytoglobin heterodimerizes with the non-histone chromatin structural protein HMGB2. Our results are consistent with a previously unknown function by which a non-erythrocytic hemoglobin inhibits DNA damage and regulates gene programs in the vasculature by modulating the genome-wide binding of HMGB2.
Keywords: Carotid artery; Cytoglobin; DNA damage; DNA repair pathways; Hemoglobin; Hydrogen peroxide; Reactive oxygen species; Redox signal; Smooth muscle.
Copyright © 2023 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest JJC is a consultant for Thermo Fisher Scientific, 908 Devices, and Seer.
Figures
Update of
-
Nuclear cytoglobin associates with HMGB2 and regulates DNA damage and genome-wide transcriptional output in the vasculature.bioRxiv [Preprint]. 2023 May 11:2023.05.10.540045. doi: 10.1101/2023.05.10.540045. bioRxiv. 2023. Update in: Redox Biol. 2023 Sep;65:102838. doi: 10.1016/j.redox.2023.102838. PMID: 37214992 Free PMC article. Updated. Preprint.
References
-
- Qiu Y., Sutton L., Riggs A.F. Identification of myoglobin in human smooth muscle. J. Biol. Chem. 1998;273:23426–23432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
