

This is a preprint.
A feedback control mechanism governs the synthesis of lipid-linked precursors of the bacterial cell wall
- PMID: 37577621
- PMCID: PMC10418202
- DOI: 10.1101/2023.08.01.551478
A feedback control mechanism governs the synthesis of lipid-linked precursors of the bacterial cell wall
Update in
-
Synthesis of lipid-linked precursors of the bacterial cell wall is governed by a feedback control mechanism in Pseudomonas aeruginosa.Nat Microbiol. 2024 Mar;9(3):763-775. doi: 10.1038/s41564-024-01603-2. Epub 2024 Feb 9. Nat Microbiol. 2024. PMID: 38336881 Free PMC article.
Abstract
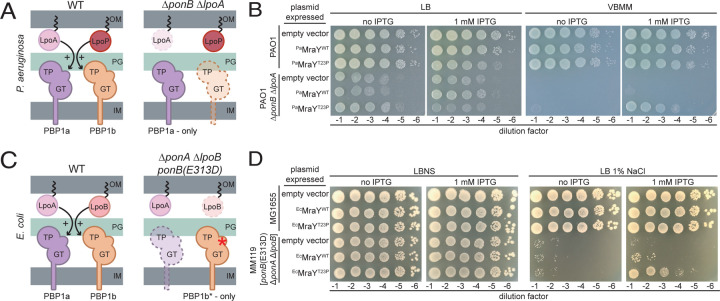
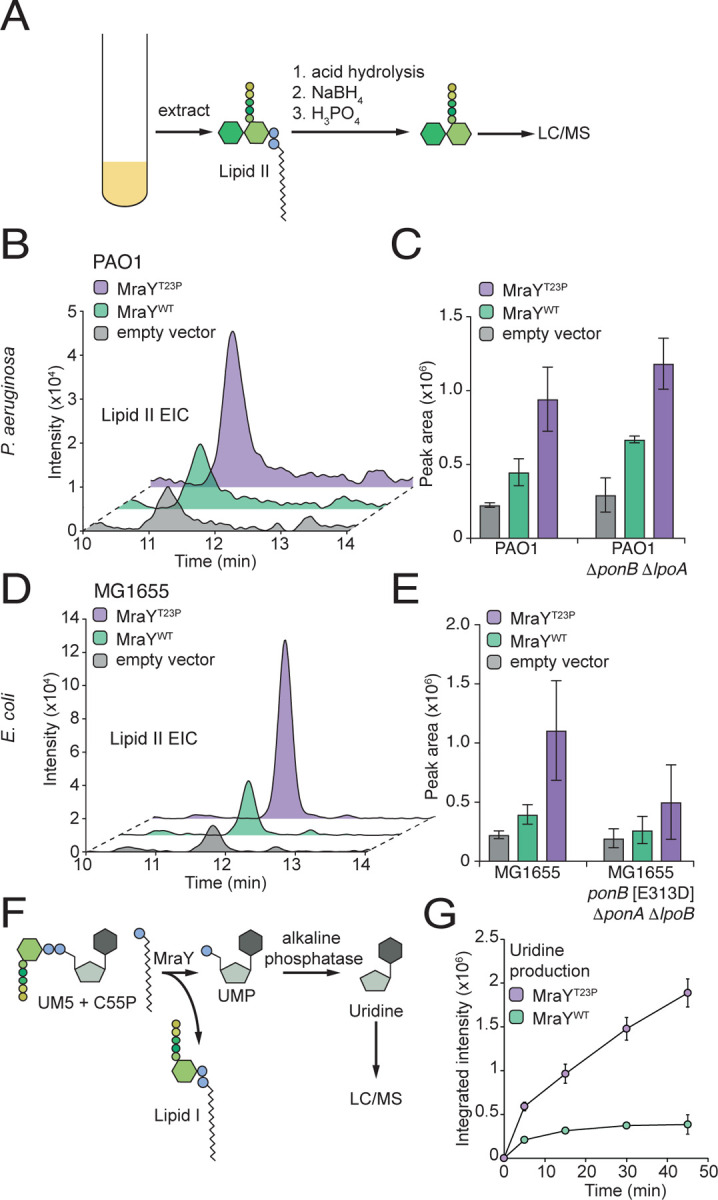
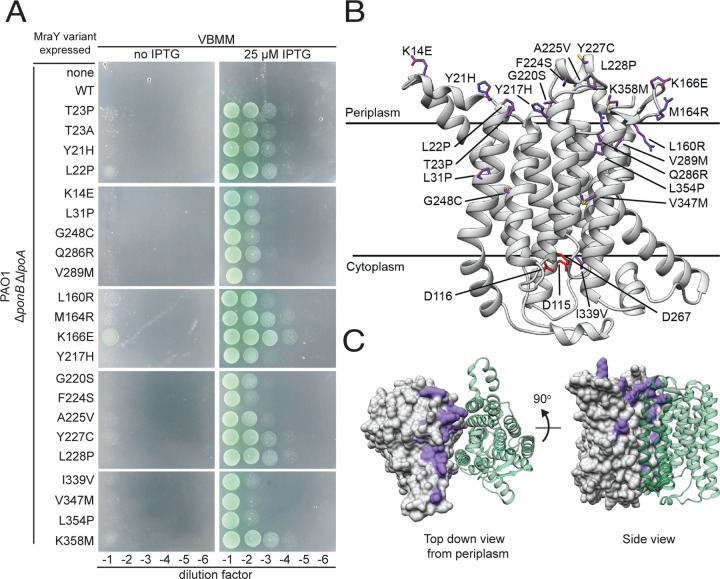
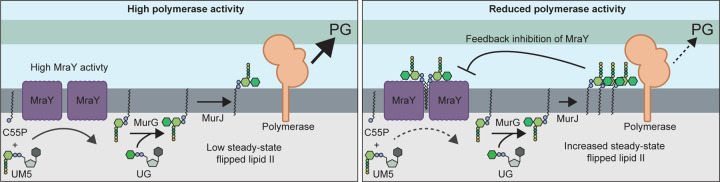
Many bacterial surface glycans such as the peptidoglycan (PG) cell wall, O-antigens, and capsules are built from monomeric units linked to a polyprenyl lipid carrier. How this limiting lipid carrier is effectively distributed among competing pathways has remained unclear for some time. Here, we describe the isolation and characterization of hyperactive variants of Pseudomonas aeruginosa MraY, the essential and conserved enzyme catalyzing the formation of the first lipid-linked PG precursor called lipid I. These variants result in the elevated production of the final PG precursor lipid II in cells and are hyperactive in a purified system. Amino acid substitutions within the activated MraY variants unexpectedly map to a cavity on the extracellular side of the dimer interface, far from the active site. Our structural evidence and molecular dynamics simulations suggest that the cavity is a binding site for lipid II molecules that have been transported to the outer leaflet of the membrane. Overall, our results support a model in which excess externalized lipid II allosterically inhibits MraY, providing a feedback mechanism to prevent the sequestration of lipid carrier in the PG biogenesis pathway. MraY belongs to the broadly distributed polyprenyl-phosphate N-acetylhexosamine 1-phosphate transferase (PNPT) superfamily of enzymes. We therefore propose that similar feedback mechanisms may be widely employed to coordinate precursor supply with demand by polymerases, thereby optimizing the partitioning of lipid carriers between competing glycan biogenesis pathways.
Keywords: Major: Biological Sciences; Minor: Microbiology; bacteriology; biochemistry; peptidoglycan; transferase.
Figures

References
-
- Rohs P. D. A. & Bernhardt T. G. Growth and Division of the Peptidoglycan Matrix. Annu. Rev. Microbiol. 75, 1–22 (2021). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources