

This is a preprint.
Molecular tuning of sea anemone stinging
- PMID: 37577638
- PMCID: PMC10418081
- DOI: 10.1101/2023.06.15.545144
Molecular tuning of sea anemone stinging
Update in
-
Molecular tuning of sea anemone stinging.Elife. 2023 Oct 31;12:RP88900. doi: 10.7554/eLife.88900. Elife. 2023. PMID: 37906220 Free PMC article.
Abstract
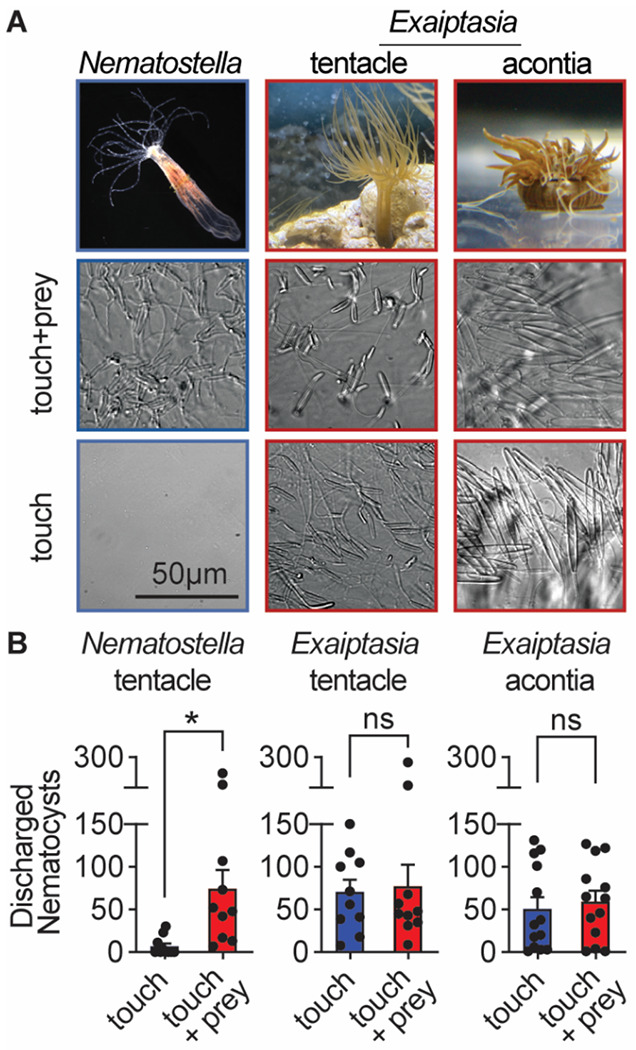
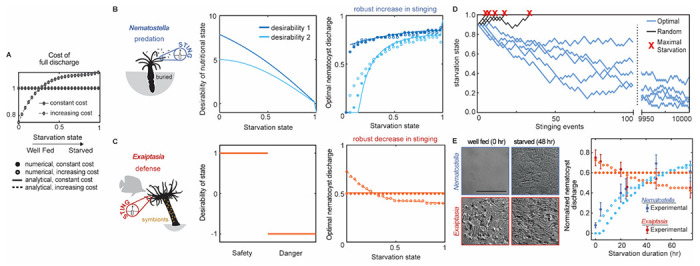
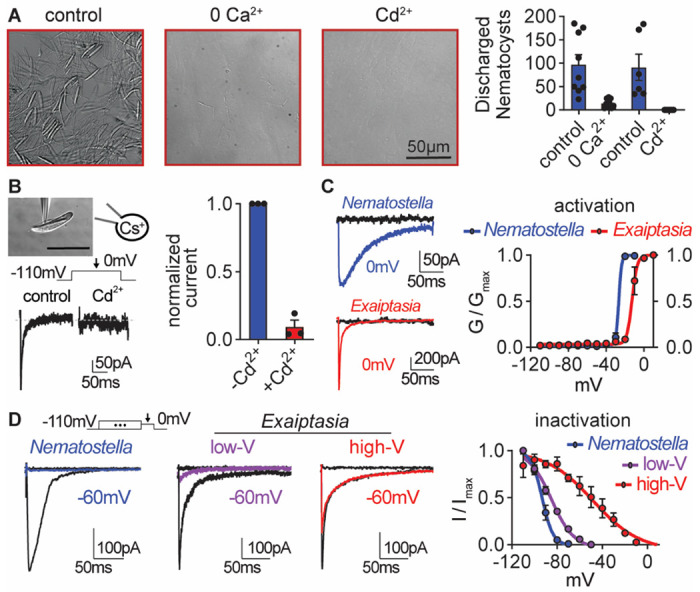
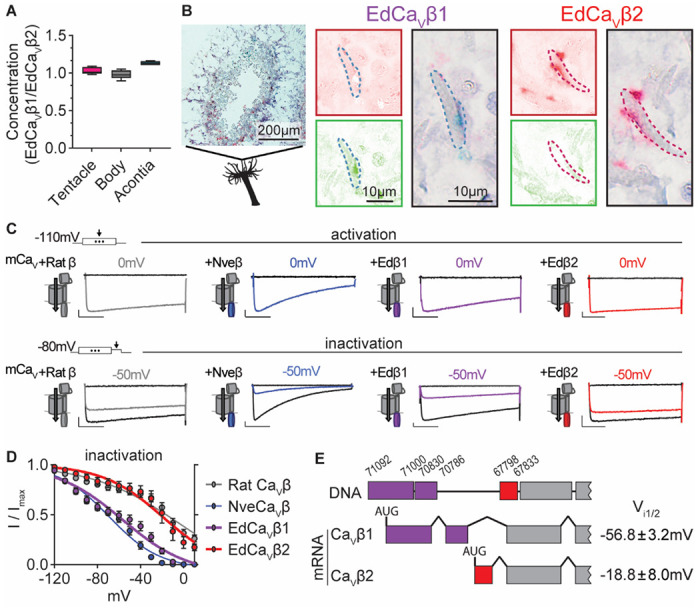
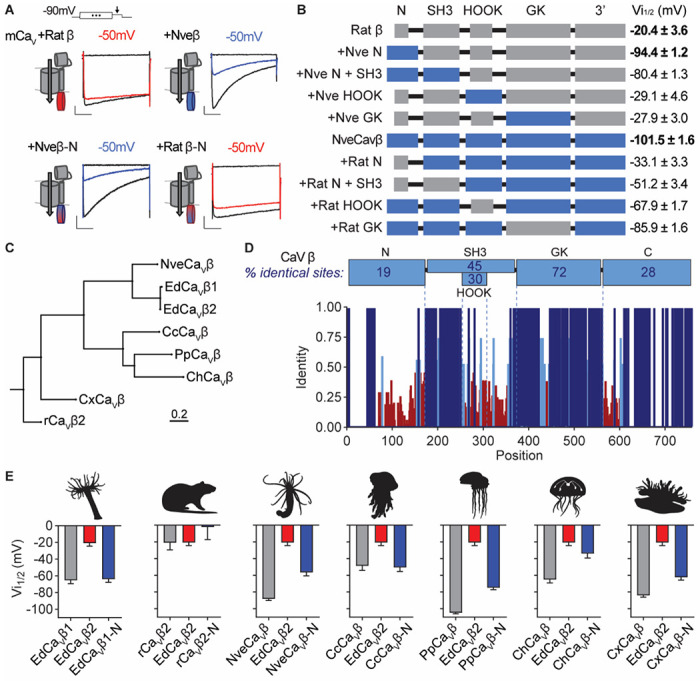
Jellyfish and sea anemones fire single-use, venom-covered barbs to immobilize prey or predators. We previously showed that the anemone Nematostella vectensis uses a specialized voltage-gated calcium (CaV) channel to trigger stinging in response to synergistic prey-derived chemicals and touch (Weir et al., 2020). Here we use experiments and theory to find that stinging behavior is suited to distinct ecological niches. We find that the burrowing anemone Nematostella uses uniquely strong CaV inactivation for precise control of predatory stinging. In contrast, the related anemone Exaiptasia diaphana inhabits exposed environments to support photosynthetic endosymbionts. Consistent with its niche, Exaiptasia indiscriminately stings for defense and expresses a CaV splice variant that confers weak inactivation. Chimeric analyses reveal that CaVβ subunit adaptations regulate inactivation, suggesting an evolutionary tuning mechanism for stinging behavior. These findings demonstrate how functional specialization of ion channel structure contributes to distinct organismal behavior.
Conflict of interest statement
Competing Interests: The authors declare no competing financial interests.
Figures
References
-
- Beckmann A, Özbek S (2012) The Nematocyst: a molecular map of the Cnidarian stinging organelle. Int J Dev Biol 56:577–582. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources