

Discontinuous L-binding motifs in the transactivation domain of the vesicular stomatitis virus P protein are required for terminal de novo transcription initiation by the L protein
- PMID: 37578231
- PMCID: PMC10506490
- DOI: 10.1128/jvi.00246-23
Discontinuous L-binding motifs in the transactivation domain of the vesicular stomatitis virus P protein are required for terminal de novo transcription initiation by the L protein
Abstract
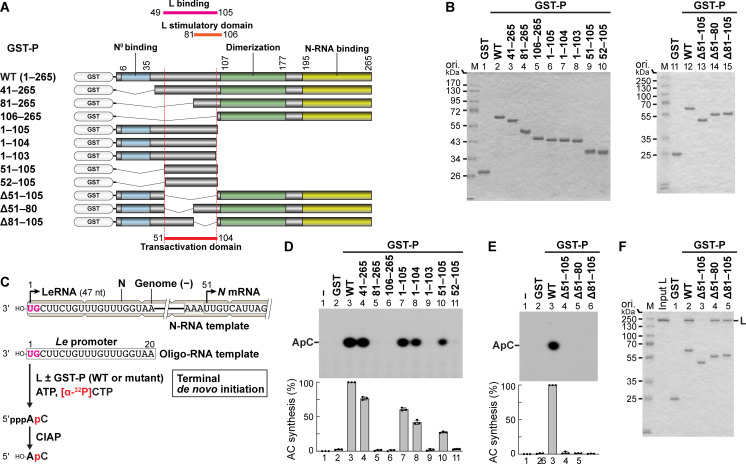
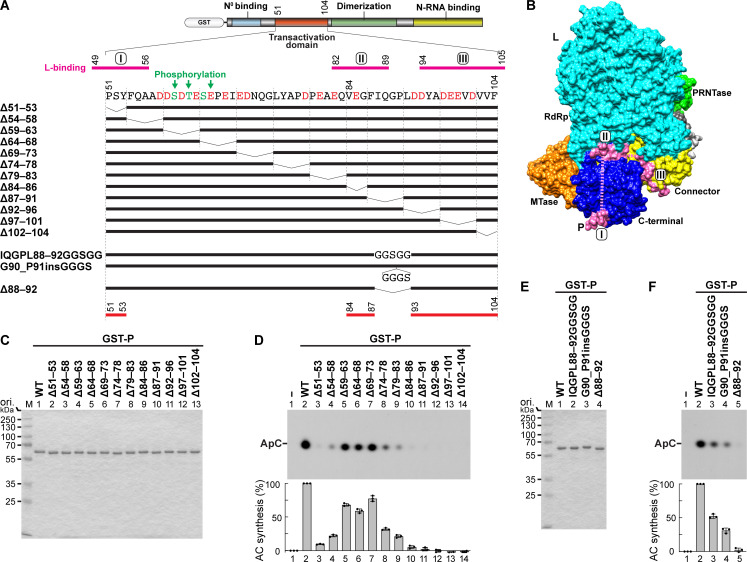
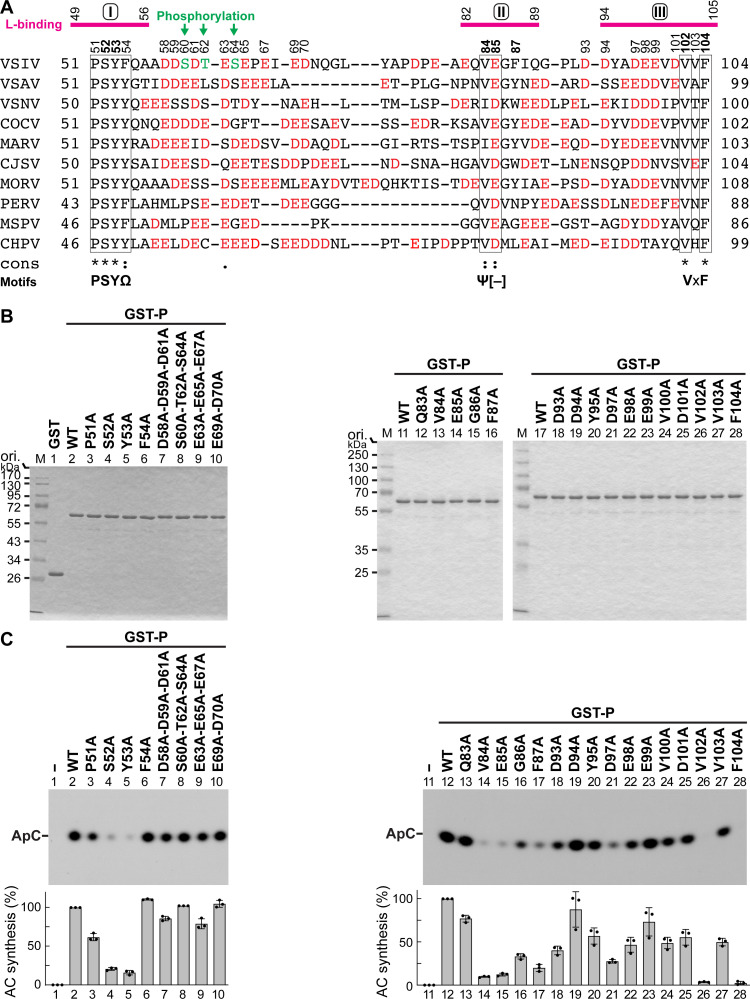
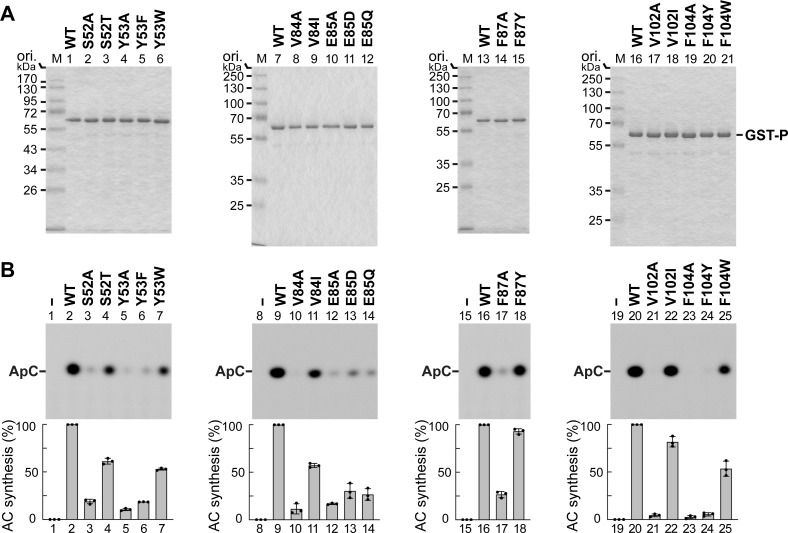
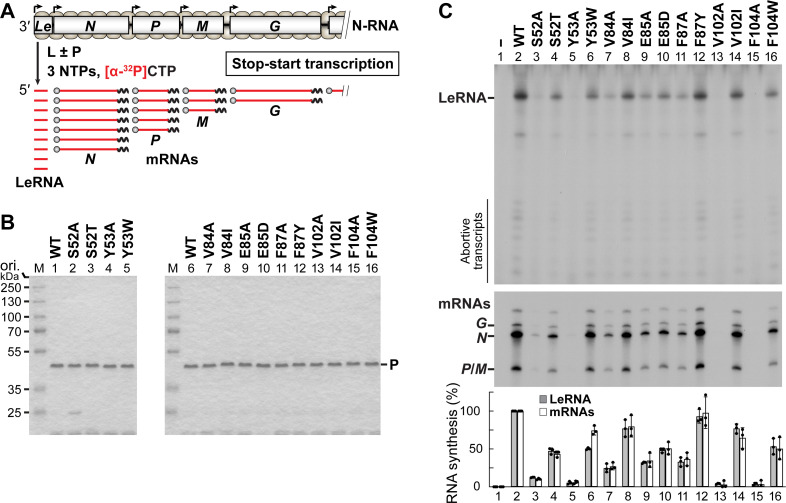
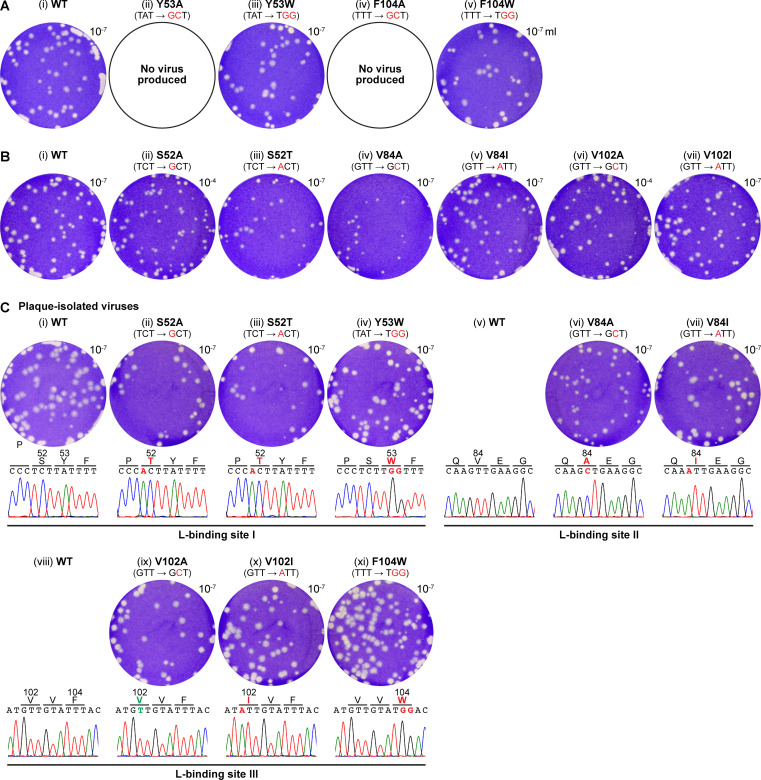
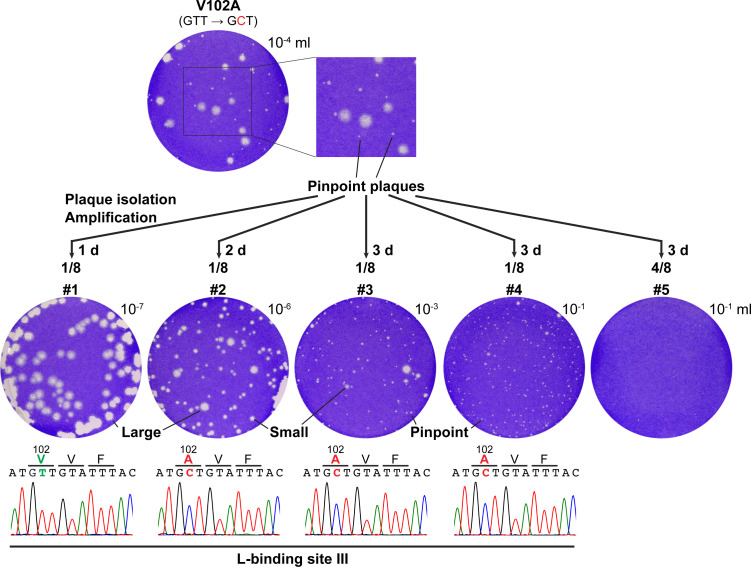
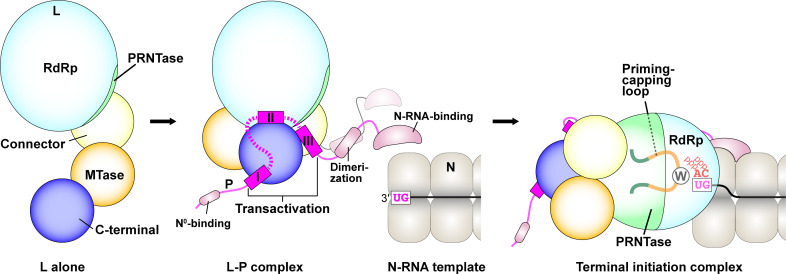
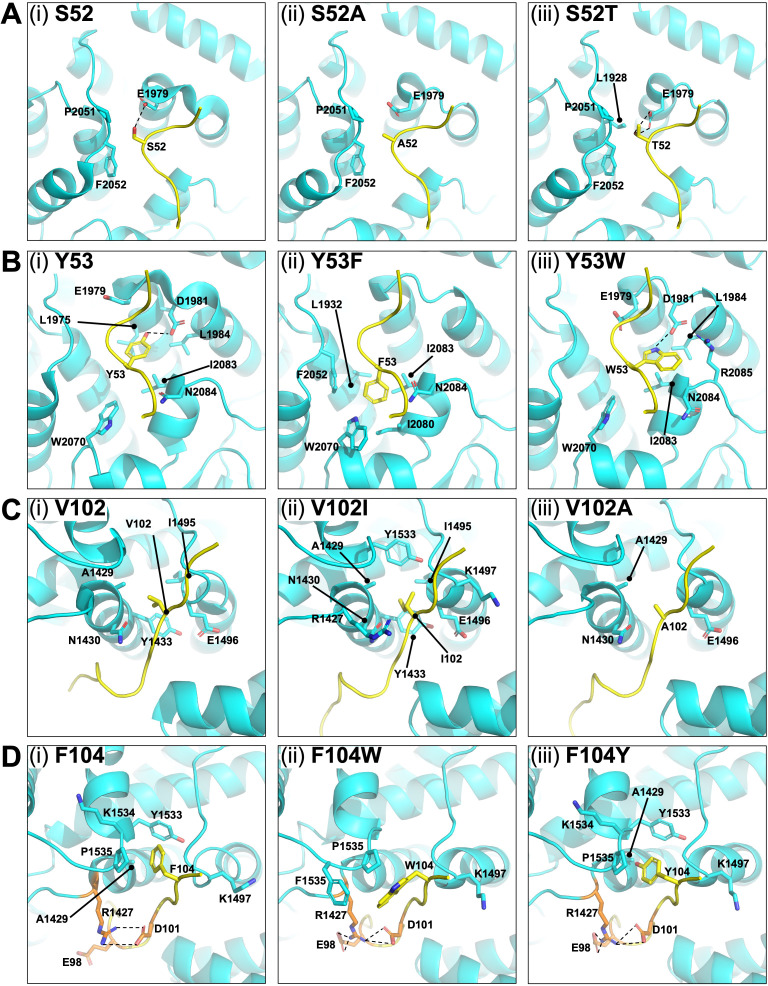
The phospho- (P) protein, the co-factor of the RNA polymerase large (L) protein, of vesicular stomatitis virus (VSV, a prototype of nonsegmented negative-strand RNA viruses) plays pivotal roles in transcription and replication. However, the precise mechanism underlying the transcriptional transactivation by the P protein has remained elusive. Here, using an in vitro transcription system and a series of deletion mutants of the P protein, we mapped a region encompassing residues 51-104 as a transactivation domain (TAD) that is critical for terminal de novo initiation, the initial step of synthesis of the leader RNA and anti-genome/genome, with the L protein. Site-directed mutagenesis revealed that conserved amino acid residues in three discontinuous L-binding sites within the TAD are essential for the transactivation activity of the P protein or important for maintaining its full activity. Importantly, relative inhibitory effects of TAD point mutations on synthesis of the full-length leader RNA and mRNAs from the 3'-terminal leader region and internal genes, respectively, of the genome were similar to those on terminal de novo initiation. Furthermore, any of the examined TAD mutations did not alter the gradient pattern of mRNAs synthesized from internal genes, nor did they induce the production of readthrough transcripts. These results suggest that these TAD mutations impact mainly terminal de novo initiation but rarely other steps (e.g., elongation, termination, internal initiation) of single-entry stop-start transcription. Consistently, the mutations of the essential or important amino acid residues within the P TAD were lethal or deleterious to VSV replication in host cells. IMPORTANCE RNA-dependent RNA polymerase L proteins of nonsegmented negative-strand RNA viruses belonging to the Mononegavirales order require their cognate co-factor P proteins or their counterparts for genome transcription and replication. However, exact roles of these co-factor proteins in modulating functions of L proteins during transcription and replication remain unknown. In this study, we revealed that three discrete L-binding motifs within a transactivation domain of the P protein of vesicular stomatitis virus, a prototypic nonsegmented negative-strand RNA virus, are required for terminal de novo initiation mediated by the L protein, which is the first step of synthesis of the leader RNA as well as genome/anti-genome.
Keywords: L protein; Mononegavirales; P protein; RNA-dependent RNA polymerase; replication; rhabdovirus; transcription; transcription factor; transcription initiation; vesicular stomatitis virus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
