

Normal and dysregulated crosstalk between iron metabolism and erythropoiesis
- PMID: 37578340
- PMCID: PMC10425177
- DOI: 10.7554/eLife.90189
Normal and dysregulated crosstalk between iron metabolism and erythropoiesis
Abstract
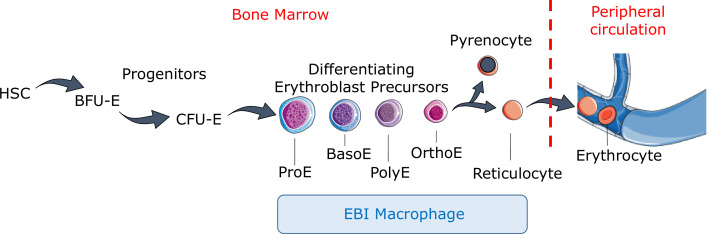
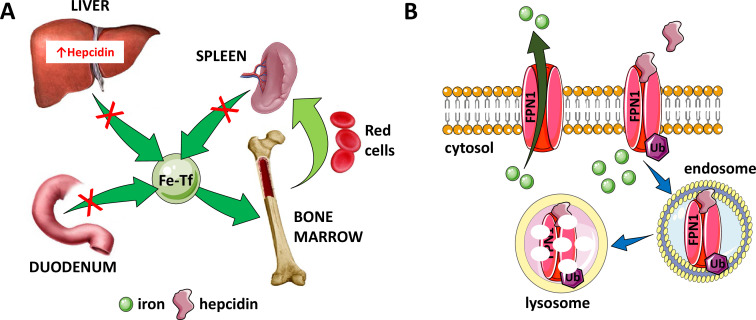
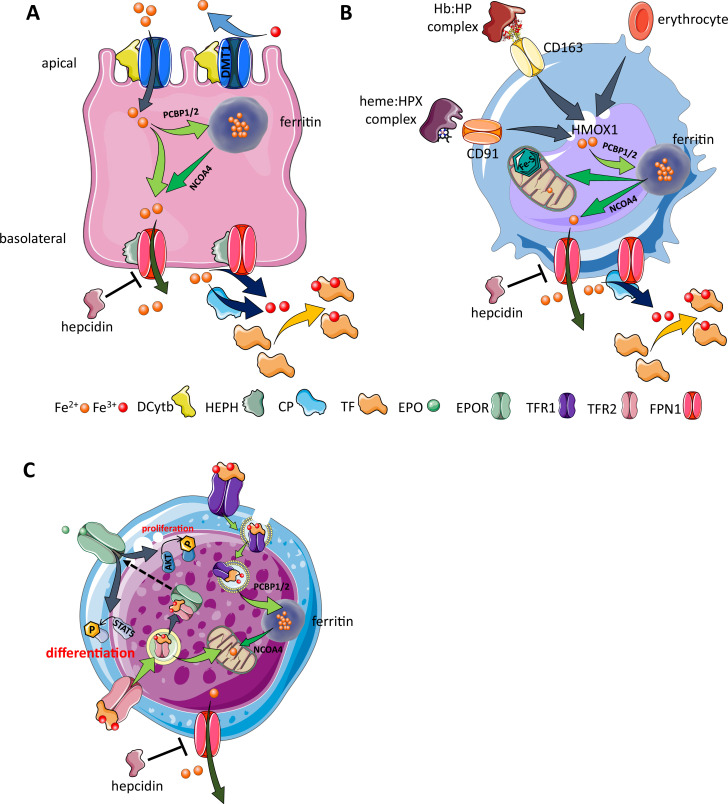
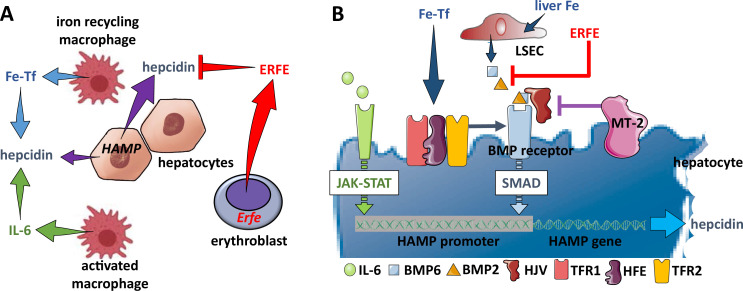
Erythroblasts possess unique characteristics as they undergo differentiation from hematopoietic stem cells. During terminal erythropoiesis, these cells incorporate large amounts of iron in order to generate hemoglobin and ultimately undergo enucleation to become mature red blood cells, ultimately delivering oxygen in the circulation. Thus, erythropoiesis is a finely tuned, multifaceted process requiring numerous properly timed physiological events to maintain efficient production of 2 million red blood cells per second in steady state. Iron is required for normal functioning in all human cells, the erythropoietic compartment consuming the majority in light of the high iron requirements for hemoglobin synthesis. Recent evidence regarding the crosstalk between erythropoiesis and iron metabolism sheds light on the regulation of iron availability by erythroblasts and the consequences of insufficient as well as excess iron on erythroid lineage proliferation and differentiation. In addition, significant progress has been made in our understanding of dysregulated iron metabolism in various congenital and acquired malignant and non-malignant diseases. Finally, we report several actual as well as theoretical opportunities for translating the recently acquired robust mechanistic understanding of iron metabolism regulation to improve management of patients with disordered erythropoiesis, such as anemia of chronic inflammation, β-thalassemia, polycythemia vera, and myelodysplastic syndromes.
Keywords: anemia; developmental biology; erythropoiesis; iron deficiency; medicine; polycythemia.
© 2023, Ginzburg et al.
Conflict of interest statement
YG Reviewing editor, eLife, XA, SR, AG No competing interests declared
Figures
Similar articles
-
β-Thalassemia and Polycythemia vera: targeting chronic stress erythropoiesis.Int J Biochem Cell Biol. 2014 Jun;51:89-92. doi: 10.1016/j.biocel.2014.03.029. Epub 2014 Apr 6. Int J Biochem Cell Biol. 2014. PMID: 24718374 Free PMC article. Review.
-
Novel potential therapeutics to modify iron metabolism and red cell synthesis in diseases associated with defective erythropoiesis.Haematologica. 2023 Oct 1;108(10):2582-2593. doi: 10.3324/haematol.2023.283057. Haematologica. 2023. PMID: 37345473 Free PMC article. Review.
-
β-thalassemia: a model for elucidating the dynamic regulation of ineffective erythropoiesis and iron metabolism.Blood. 2011 Oct 20;118(16):4321-30. doi: 10.1182/blood-2011-03-283614. Epub 2011 Jul 18. Blood. 2011. PMID: 21768301 Free PMC article. Review.
-
Macrophages support pathological erythropoiesis in polycythemia vera and β-thalassemia.Nat Med. 2013 Apr;19(4):437-45. doi: 10.1038/nm.3126. Epub 2013 Mar 17. Nat Med. 2013. PMID: 23502961 Free PMC article.
-
Hemoglobinization and functional availability of iron for erythropoiesis in case of thalassemia and iron deficiency anemia.Clin Lab. 2006;52(3-4):107-14. Clin Lab. 2006. PMID: 16584056
Cited by
-
Endogenous small molecule effectors in GATA transcription factor mechanisms governing biological and pathological processes.Exp Hematol. 2024 Sep;137:104252. doi: 10.1016/j.exphem.2024.104252. Epub 2024 Jun 12. Exp Hematol. 2024. PMID: 38876253 Review.
-
Improved MRI detection of inflammation-induced changes in bone marrow microstructure in mice: a machine learning-enhanced T2 distribution analysis.Eur Radiol Exp. 2025 Mar 26;9(1):39. doi: 10.1186/s41747-025-00574-1. Eur Radiol Exp. 2025. PMID: 40140174 Free PMC article.
-
PLK1 inhibition impairs erythroid differentiation.Front Cell Dev Biol. 2024 Dec 23;12:1516704. doi: 10.3389/fcell.2024.1516704. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39763588 Free PMC article.
-
Diagnostics: Markers of Body Iron Status.Adv Exp Med Biol. 2025;1480:387-398. doi: 10.1007/978-3-031-92033-2_25. Adv Exp Med Biol. 2025. PMID: 40603804 Review.
-
Utility of reticulocyte hemoglobin as a new predictor of anemia in intensive care unit patients.Front Med (Lausanne). 2025 Apr 9;12:1577047. doi: 10.3389/fmed.2025.1577047. eCollection 2025. Front Med (Lausanne). 2025. PMID: 40270491 Free PMC article.
References
-
- Adema V, Ma F, Kanagal-Shamanna R, Thongon N, Montalban-Bravo G, Yang H, Peslak SA, Wang F, Acha P, Sole F, Lockyer P, Cassari M, Maciejewski JP, Visconte V, Gañán-Gómez I, Song Y, Bueso-Ramos C, Pellegrini M, Tan TM, Bejar R, Carew JS, Halene S, Santini V, Al-Atrash G, Clise-Dwyer K, Garcia-Manero G, Blobel GA, Colla S. Targeting the EIF2AK1 signaling pathway rescues red blood cell production in SF3B1-mutant myelodysplastic syndromes with ringed sideroblasts. Blood Cancer Discovery. 2022;3:554–567. doi: 10.1158/2643-3230.BCD-21-0220. - DOI - PMC - PubMed
-
- Ali AM, Huang Y, Pinheiro RF, Xue F, Hu J, Iverson N, Hoehn D, Coutinho D, Kayani J, Chernak B, Lane J, Hillyer C, Galili N, Jurcic J, Mohandas N, An X, Raza A. Severely impaired terminal erythroid differentiation as an independent prognostic marker in myelodysplastic syndromes. Blood Advances. 2018;2:1393–1402. doi: 10.1182/bloodadvances.2018018440. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL140625/HL/NHLBI NIH HHS/United States
- DK133475/NH/NIH HHS/United States
- DK079924/NH/NIH HHS/United States
- R56 DK132146/DK/NIDDK NIH HHS/United States
- DK107670/NH/NIH HHS/United States
- DK101550/NH/NIH HHS/United States
- R01 DK107670/DK/NIDDK NIH HHS/United States
- DK132146/NH/NIH HHS/United States
- R01 HL165202/HL/NHLBI NIH HHS/United States
- R01 DK079924/DK/NIDDK NIH HHS/United States
- HL149626/NH/NIH HHS/United States
- R01 DK133475/DK/NIDDK NIH HHS/United States
- P01 HL149626/HL/NHLBI NIH HHS/United States
- R56 HL165202/HL/NHLBI NIH HHS/United States
- R01 DK095112/DK/NIDDK NIH HHS/United States
- HL140625/NH/NIH HHS/United States
- HL165202/NH/NIH HHS/United States
- DK095112/NH/NIH HHS/United States
- R01 DK101550/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
