

Crosstalk between glucocorticoid and mineralocorticoid receptors boosts glucocorticoid-induced killing of multiple myeloma cells
- PMID: 37578563
- PMCID: PMC10425521
- DOI: 10.1007/s00018-023-04900-x
Crosstalk between glucocorticoid and mineralocorticoid receptors boosts glucocorticoid-induced killing of multiple myeloma cells
Abstract
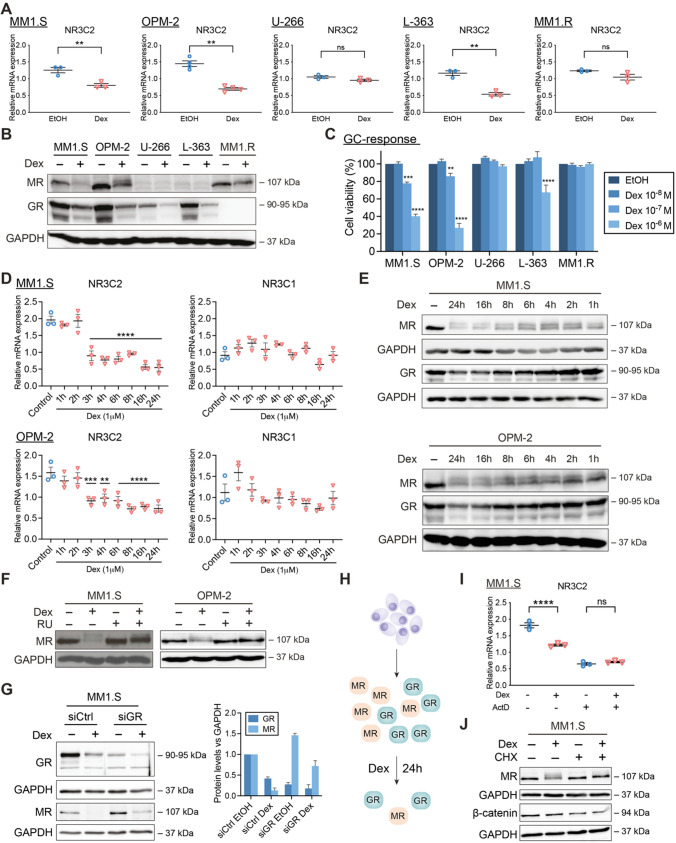
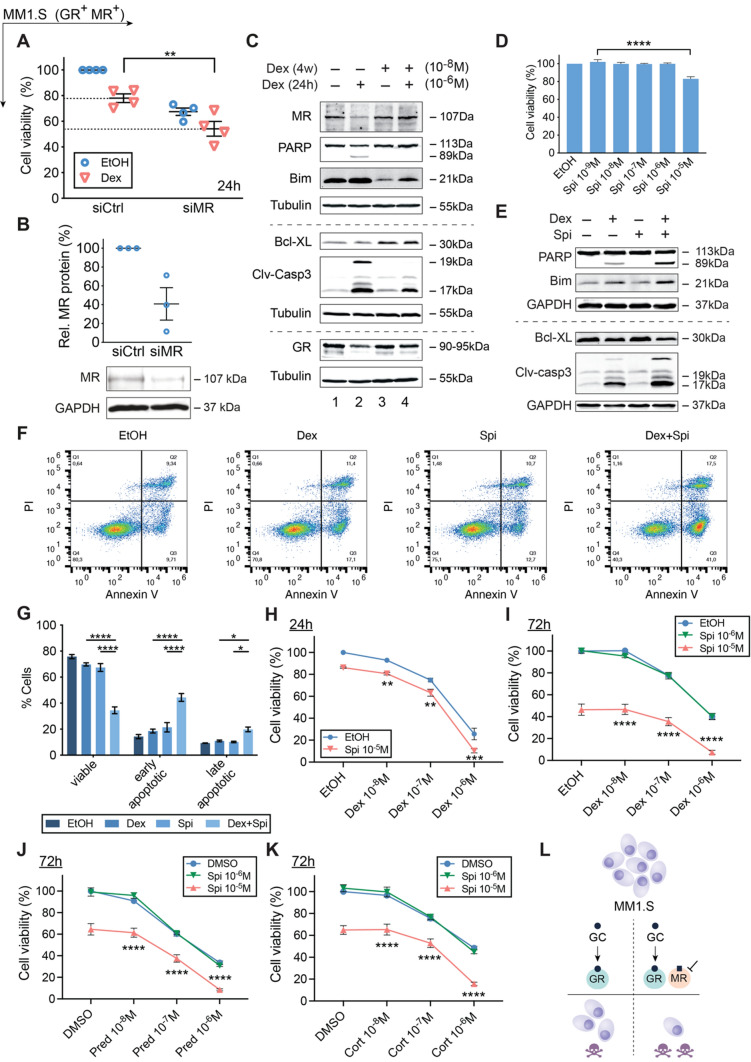
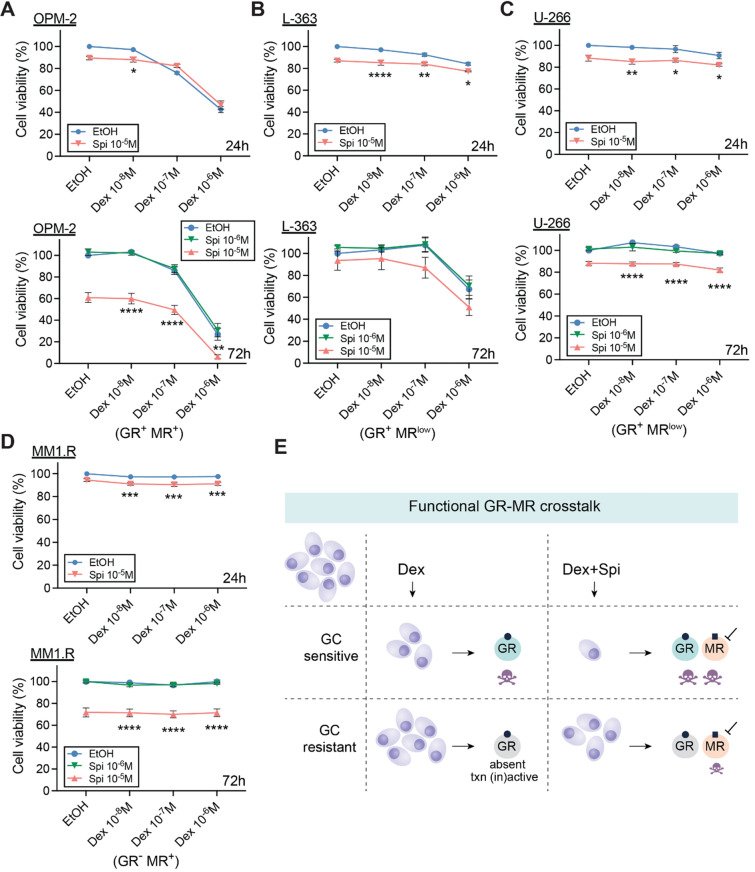
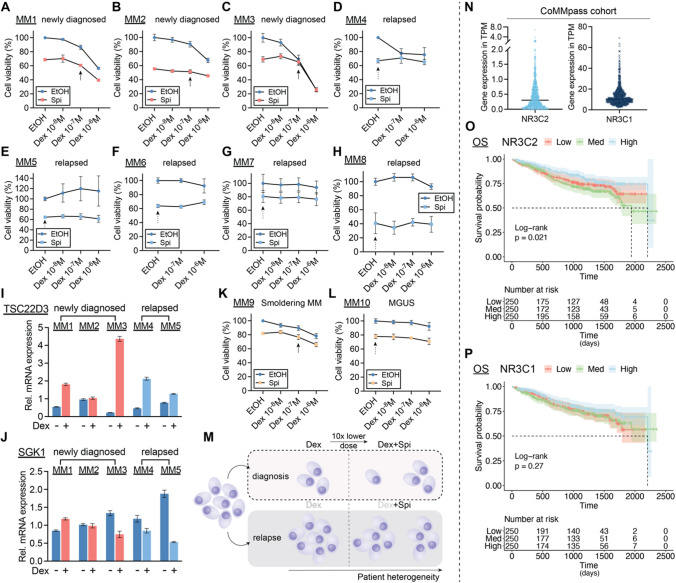
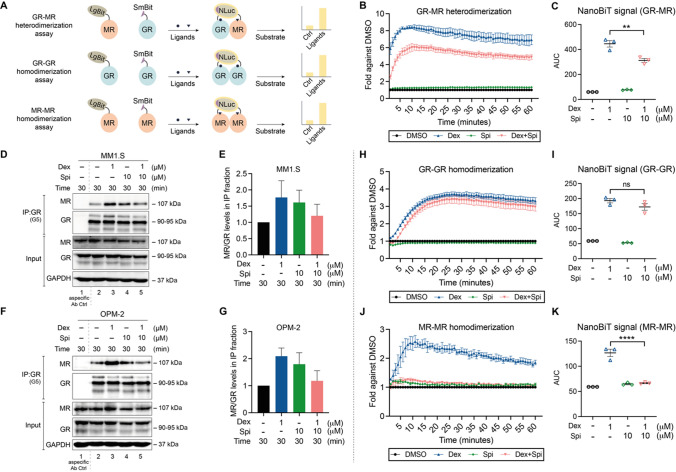
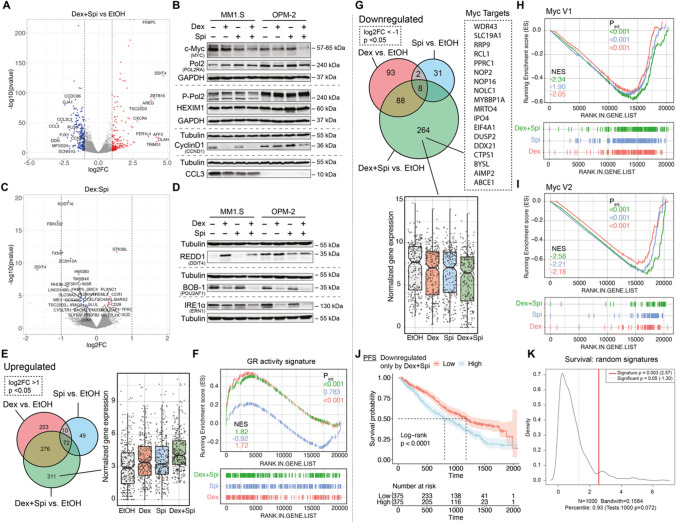
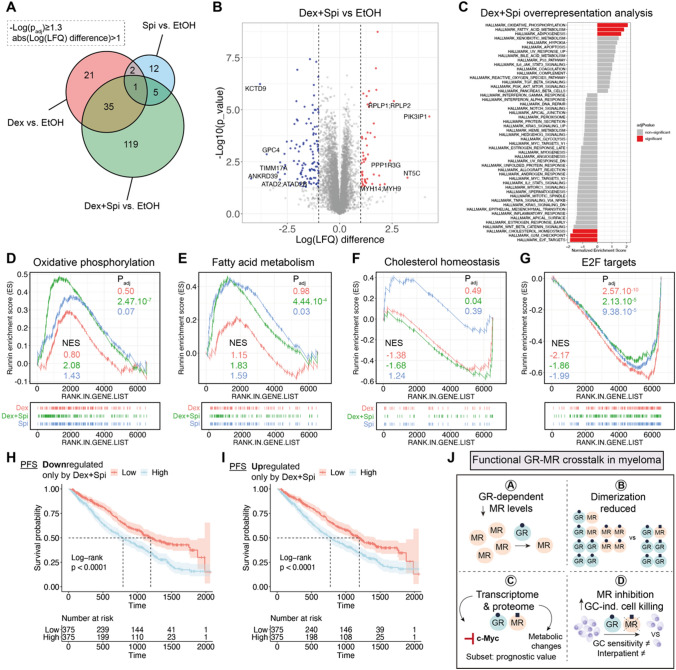
The glucocorticoid receptor (GR) is a crucial drug target in multiple myeloma as its activation with glucocorticoids effectively triggers myeloma cell death. However, as high-dose glucocorticoids are also associated with deleterious side effects, novel approaches are urgently needed to improve GR action in myeloma. Here, we reveal a functional crosstalk between GR and the mineralocorticoid receptor (MR) that plays a role in improved myeloma cell killing. We show that the GR agonist dexamethasone (Dex) downregulates MR levels in a GR-dependent way in myeloma cells. Co-treatment of Dex with the MR antagonist spironolactone (Spi) enhances Dex-induced cell killing in primary, newly diagnosed GC-sensitive myeloma cells. In a relapsed GC-resistant setting, Spi alone induces distinct myeloma cell killing. On a mechanistic level, we find that a GR-MR crosstalk likely arises from an endogenous interaction between GR and MR in myeloma cells. Quantitative dimerization assays show that Spi reduces Dex-induced GR-MR heterodimerization and completely abolishes Dex-induced MR-MR homodimerization, while leaving GR-GR homodimerization intact. Unbiased transcriptomics analyses reveal that c-myc and many of its target genes are downregulated most by combined Dex-Spi treatment. Proteomics analyses further identify that several metabolic hallmarks are modulated most by this combination treatment. Finally, we identified a subset of Dex-Spi downregulated genes and proteins that may predict prognosis in the CoMMpass myeloma patient cohort. Our study demonstrates that GR-MR crosstalk is therapeutically relevant in myeloma as it provides novel strategies for glucocorticoid-based dose-reduction.
Keywords: Glucocorticoid receptor; Glucocorticoids; Mineralocorticoid receptor; Multiple myeloma; Nuclear receptor crosstalk.
© 2023. The Author(s).
Conflict of interest statement
The authors have no relevant financial or non-financial interests to disclose.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
