

Ceramide as an endothelial cell surface receptor and a lung-specific lipid vascular target for circulating ligands
- PMID: 37579172
- PMCID: PMC10450669
- DOI: 10.1073/pnas.2220269120
Ceramide as an endothelial cell surface receptor and a lung-specific lipid vascular target for circulating ligands
Abstract
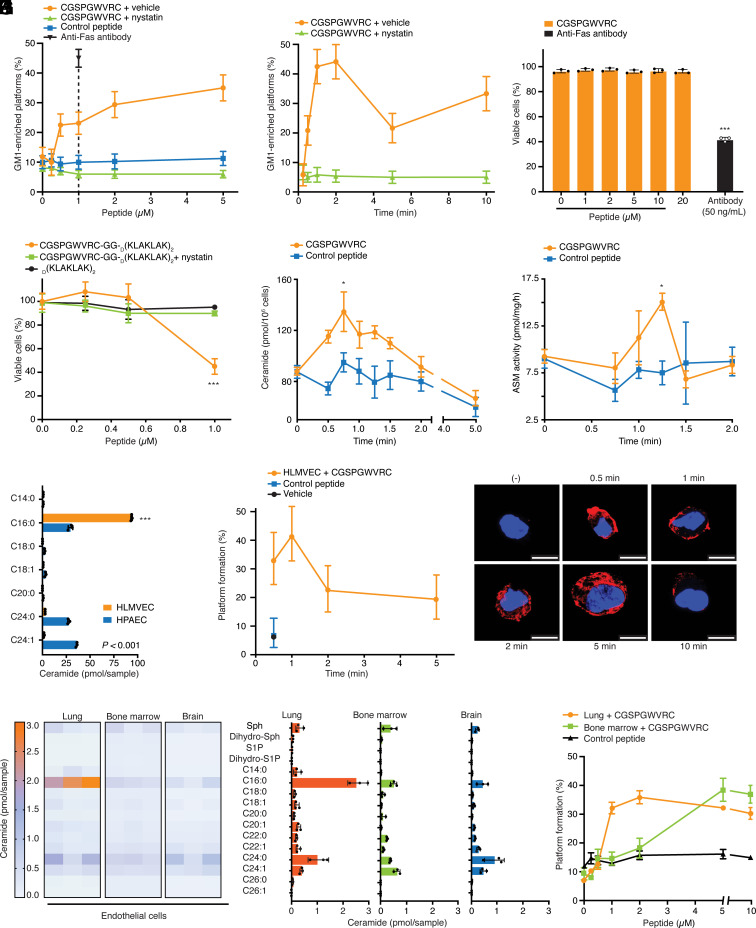
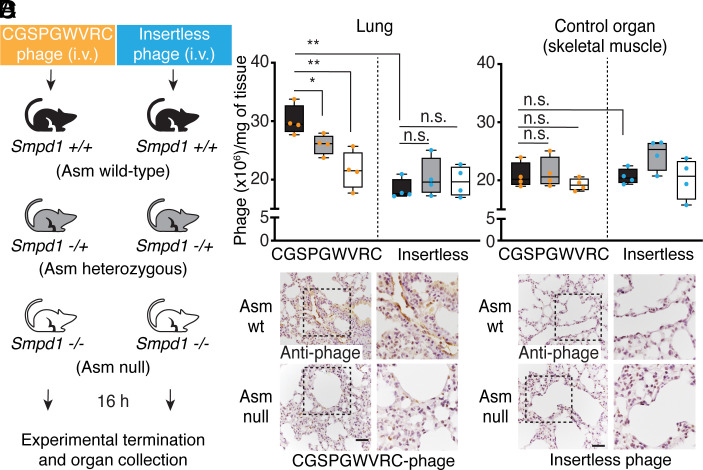
The vascular endothelium from individual organs is functionally specialized, and it displays a unique set of accessible molecular targets. These serve as endothelial cell receptors to affinity ligands. To date, all identified vascular receptors have been proteins. Here, we show that an endothelial lung-homing peptide (CGSPGWVRC) interacts with C16-ceramide, a bioactive sphingolipid that mediates several biological functions. Upon binding to cell surfaces, CGSPGWVRC triggers ceramide-rich platform formation, activates acid sphingomyelinase and ceramide production, without the associated downstream apoptotic signaling. We also show that the lung selectivity of CGSPGWVRC homing peptide is dependent on ceramide production in vivo. Finally, we demonstrate two potential applications for this lipid vascular targeting system: i) as a bioinorganic hydrogel for pulmonary imaging and ii) as a ligand-directed lung immunization tool against COVID-19. Thus, C16-ceramide is a unique example of a lipid-based receptor system in the lung vascular endothelium targeted in vivo by circulating ligands such as CGSPGWVRC.
Keywords: acid sphingomyelinase; ceramide; endothelial cells; lung; phage display.
Conflict of interest statement
J.A.R., R.N.K., R.P., and W.A. are inventors on US patents #8,562,993, #9,592,238 and #11,207,329, which describe the anti-ceramide monoclonal antibody 2A2 used in this study; as such, they will be entitled to standard royalties if commercialization occurs. J.G.G., R.P., and W.A. are founders and equity stockholders of PhageNova Bio. R.P. and W.A. receive research support from PhageNova Bio. R.P. is a paid consultant for PhageNova Bio and serves as its Chief Scientific Officer. D.I.S., F.H.F.T., T.L.S., R.P., and W.A. are inventors on international patent application PCT/US2021/040392, which describes immunization technology adapted for COVID-19. PhageNova Bio has licensed this intellectual property portfolio and D.I.S., F.H.F.T., T.L.S., R.P., and W.A. will be entitled to standard royalties if commercialization occurs. R.P. and W.A. are founders, stockholders, and scientific advisors for MBrace Therapeutics, a company developing antibodies for oncology applications. R.P. and W.A. receive research support from MBrace Therapeutics. R.P. and W.A. are paid consultants for MBrace Therapeutics. R.N.K. is founder and stockholder of Ceramedix Holding LLC. R.N.K. is an inventor on US patent #10,413,533 and pending international patent applications describing methods to activate the ceramide/sphingomyelin pathway; he will be entitled to standard royalties if commercialization occurs. J.A.R. and R.N.K. are inventors on US patent #10,450,385 and pending US and international patent applications describing anti-ceramide antibodies; as such, they will be entitled to standard royalties if commercialization occurs. R.N.K. is an inventor on US patents #7,195,775 and #7,850,984 describing methods of use of antagonists to acid sphingomyelinase and US patent #10,052,387 describing methods of use of exogenous ceramide for therapy. Y.A.H. and L.M.O. are inventors on US patent #8,093,393. Y.A.H. is also inventor on US patents #8,592,419 and #8,697,379, all of which describe ceramide-related compounds; as such, they will be entitled to standard royalties if commercialization occurs. Each of the corresponding institutions currently manages these arrangements according to their established institutional conflict-of-interest policies. None of these conflicts affected the experimental design, interpretation, or reporting of the results. Other authors declare that they have no competing interests.
Figures



References
-
- Pasqualini R., Ruoslahti E., Organ targeting in vivo using phage display peptide libraries. Nature 380, 364–366 (1996). - PubMed
-
- Arap W., et al. , Steps toward mapping the human vasculature by phage display. Nat. Med. 8, 121–127 (2002). - PubMed
-
- Arap W., Pasqualini R., Ruoslahti E., Cancer treatment by targeted drug delivery to tumor vasculature in a mouse model. Science 279, 377–380 (1998). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
