

Toll-like receptor 4 and macrophage scavenger receptor 1 crosstalk regulates phagocytosis of a fungal pathogen
- PMID: 37580395
- PMCID: PMC10425417
- DOI: 10.1038/s41467-023-40635-w
Toll-like receptor 4 and macrophage scavenger receptor 1 crosstalk regulates phagocytosis of a fungal pathogen
Abstract
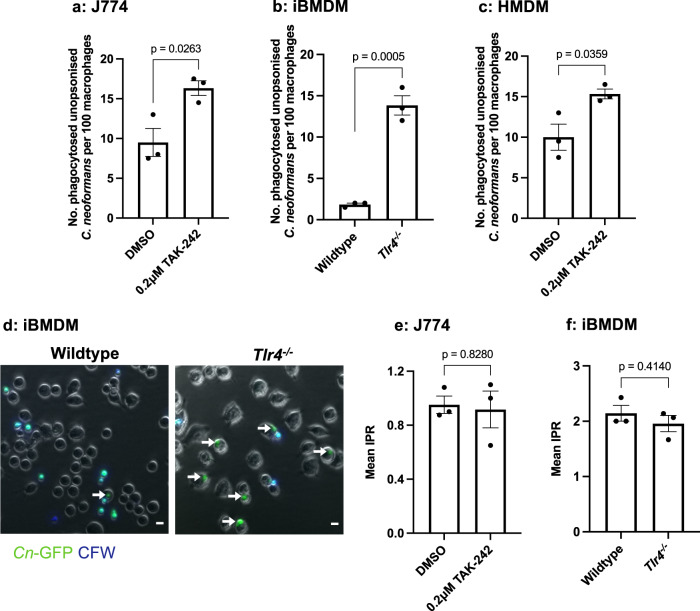
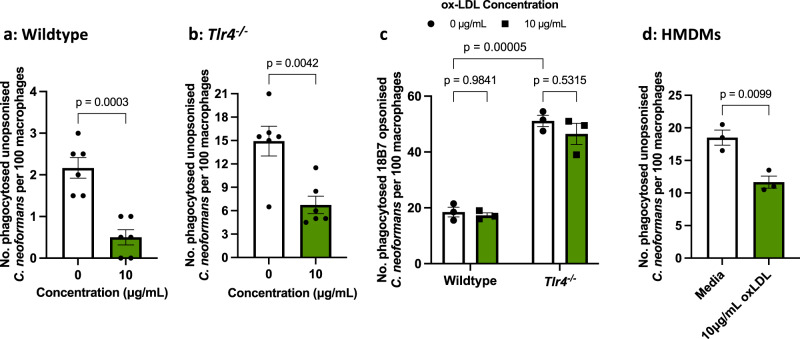
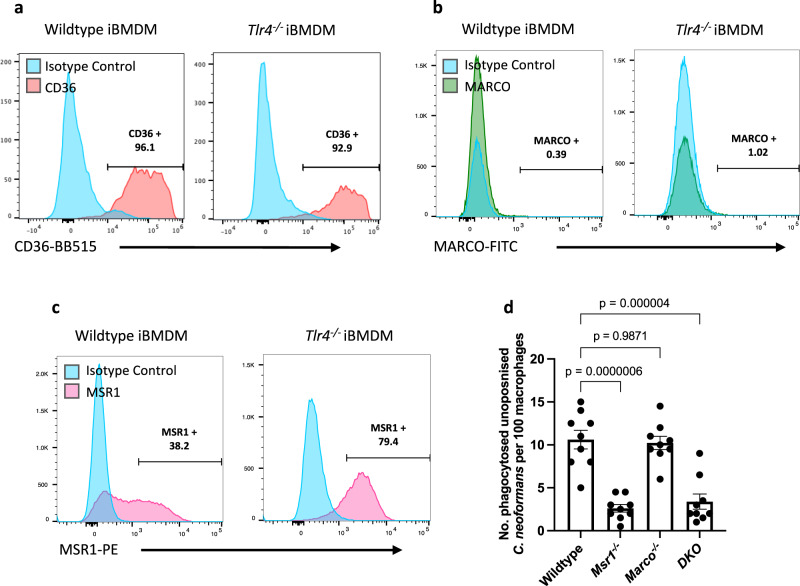
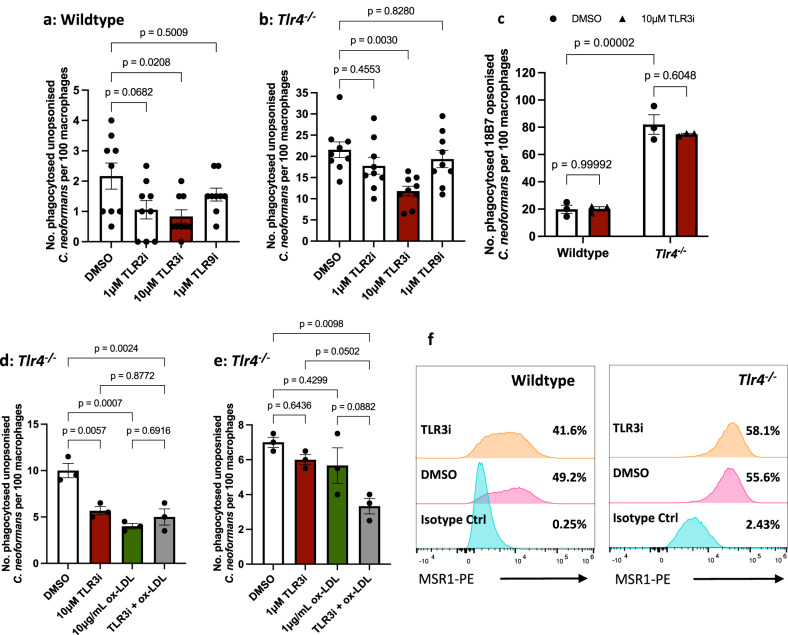
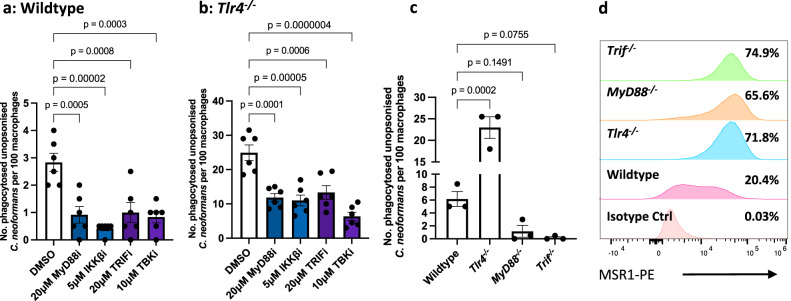
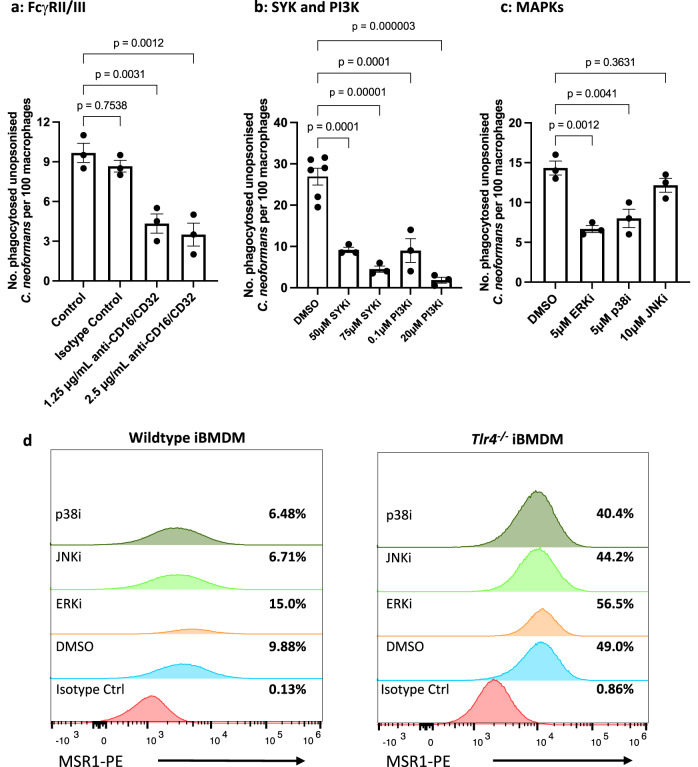
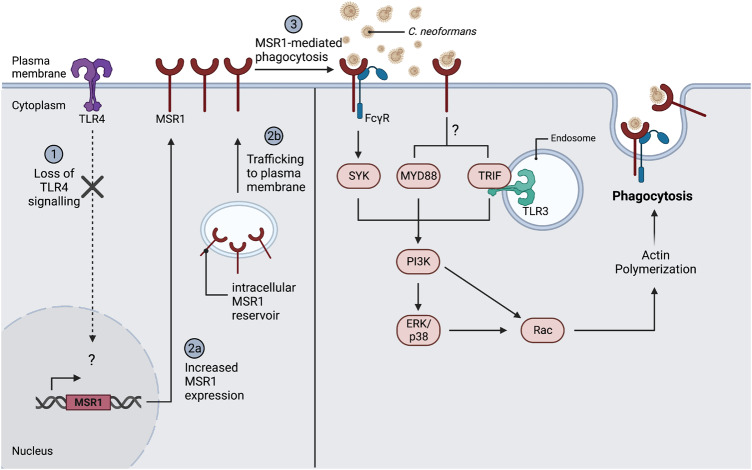
The opportunistic fungal pathogen Cryptococcus neoformans causes lethal infections in immunocompromised patients. Macrophages are central to the host response to cryptococci; however, it is unclear how C. neoformans is recognised and phagocytosed by macrophages. Here we investigate the role of TLR4 in the non-opsonic phagocytosis of C. neoformans. We find that loss of TLR4 function unexpectedly increases phagocytosis of non-opsonised cryptococci by murine and human macrophages. The increased phagocytosis observed in Tlr4-/- cells was dampened by pre-treatment of macrophages with oxidised-LDL, a known ligand of scavenger receptors. The scavenger receptor, macrophage scavenger receptor 1 (MSR1) (also known as SR-A1 or CD204) was upregulated in Tlr4-/- macrophages. Genetic ablation of MSR1 resulted in a 75% decrease in phagocytosis of non-opsonised cryptococci, strongly suggesting that it is a key non-opsonic receptor for this pathogen. We go on to show that MSR1-mediated uptake likely involves the formation of a multimolecular signalling complex involving FcγR leading to SYK, PI3K, p38 and ERK1/2 activation to drive actin remodelling and phagocytosis. Altogether, our data indicate a hitherto unidentified role for TLR4/MSR1 crosstalk in the non-opsonic phagocytosis of C. neoformans.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
