

NAD+ salvage governs the immunosuppressive capacity of mesenchymal stem cells
- PMID: 37580400
- PMCID: PMC10541442
- DOI: 10.1038/s41423-023-01073-2
NAD+ salvage governs the immunosuppressive capacity of mesenchymal stem cells
Abstract
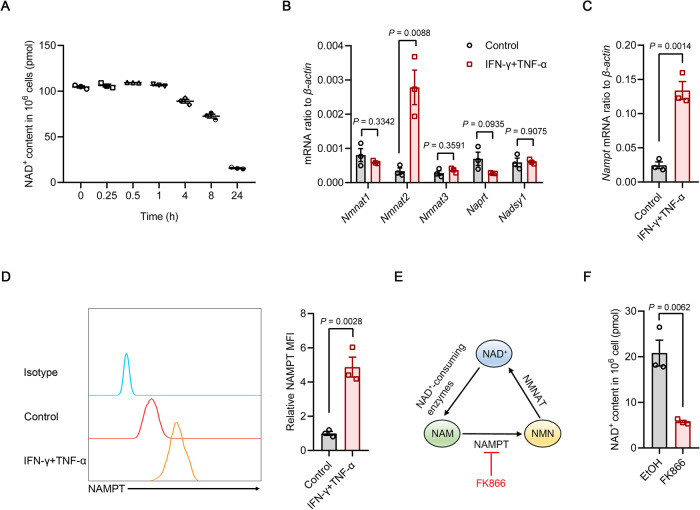
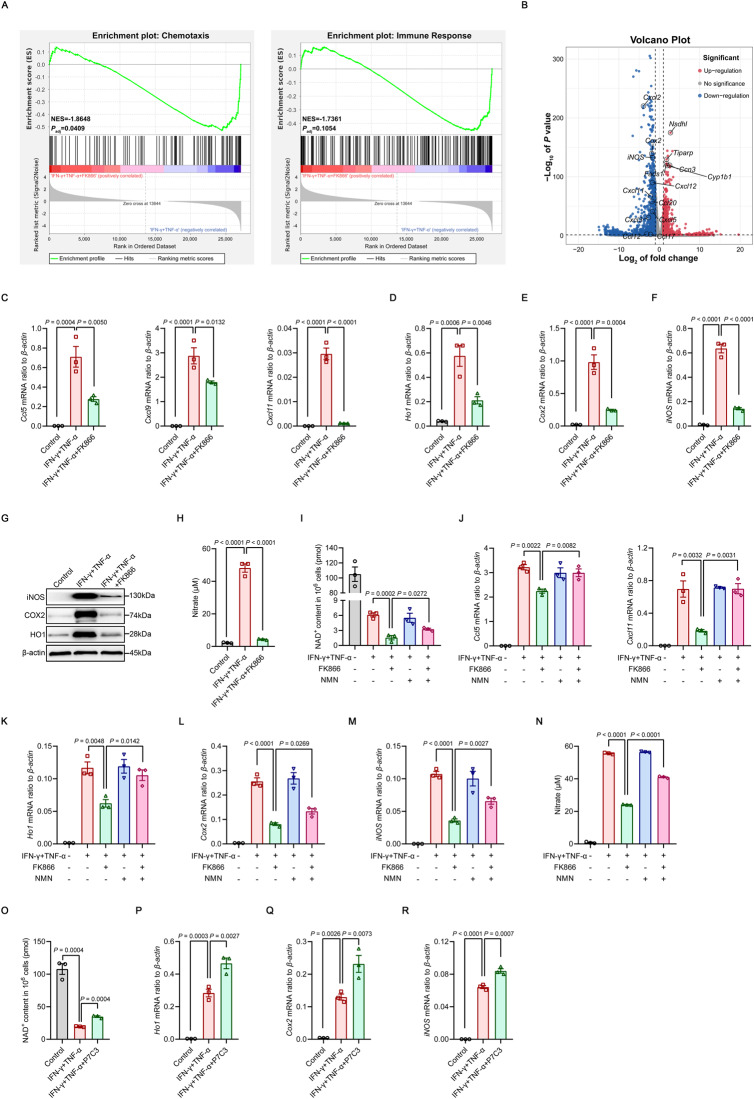
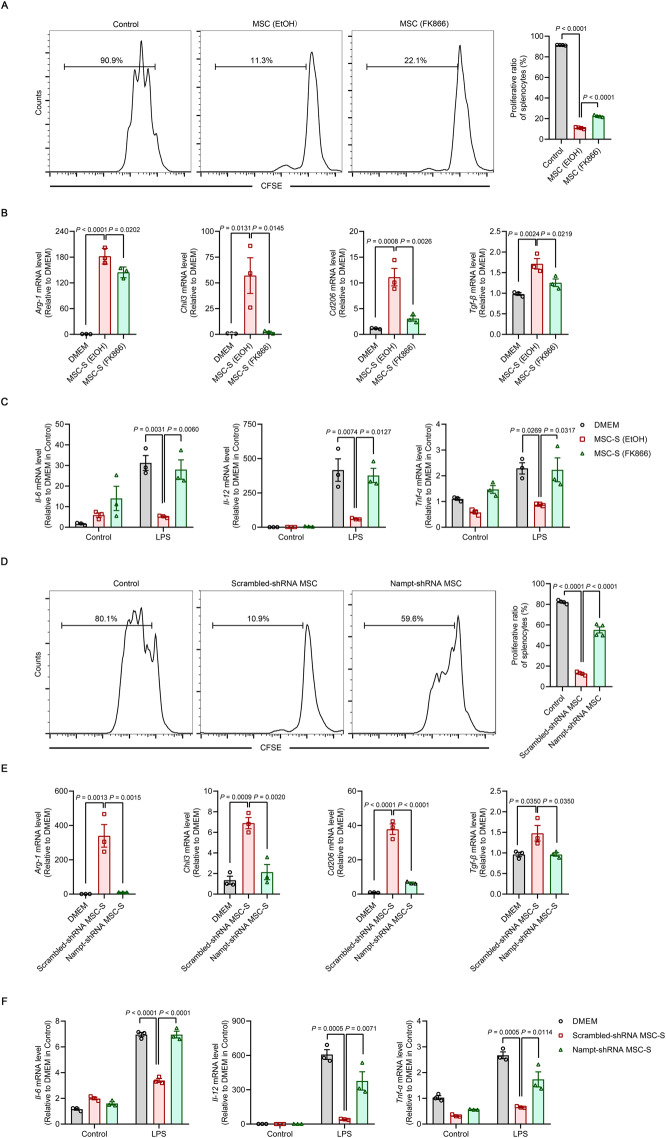
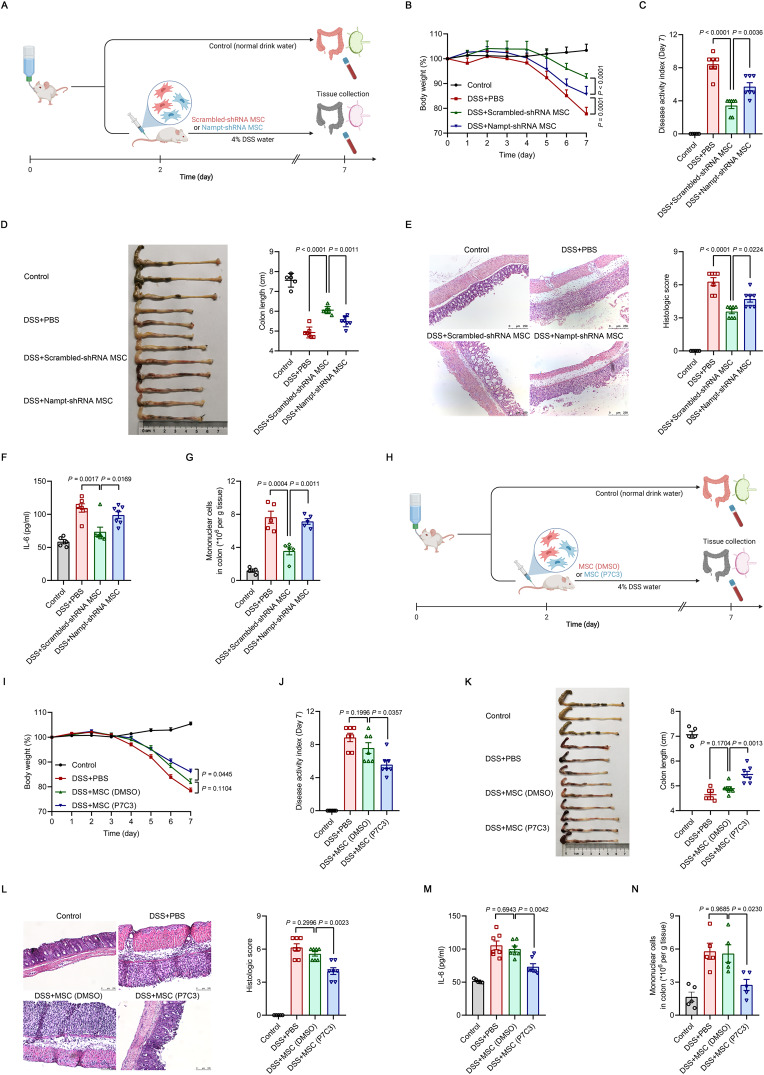
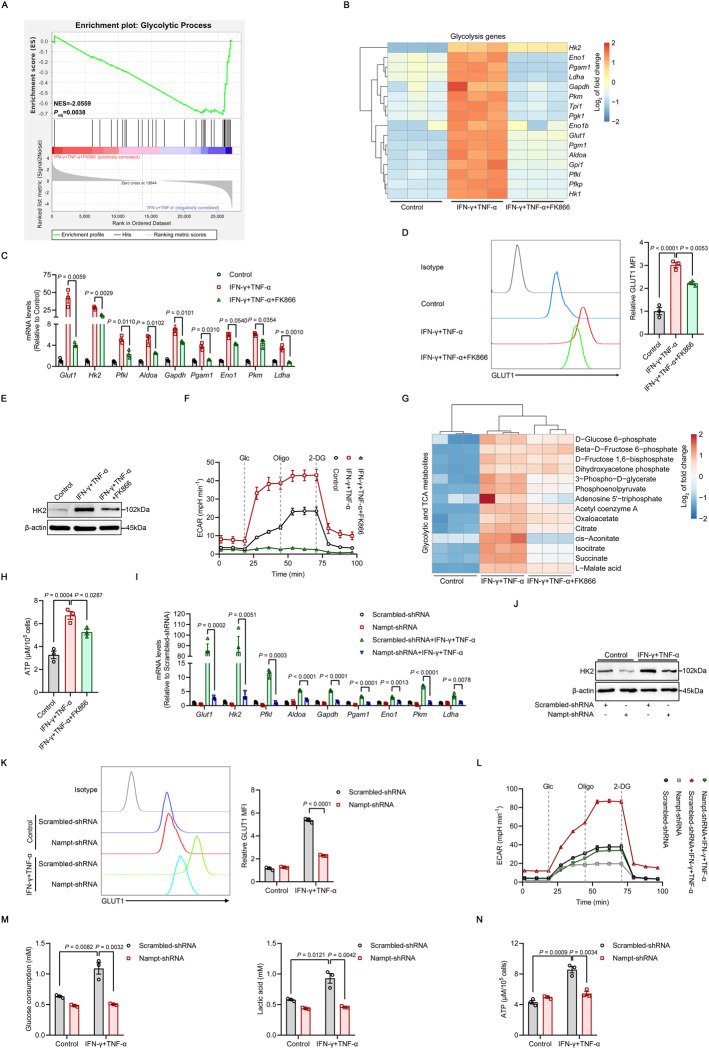
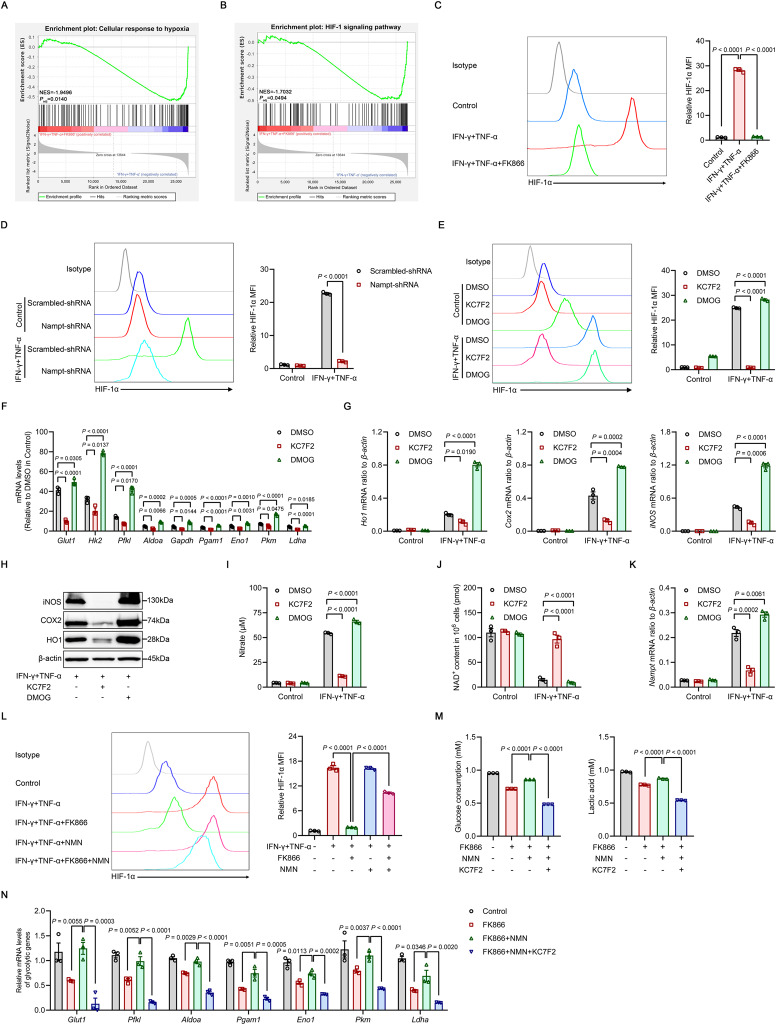
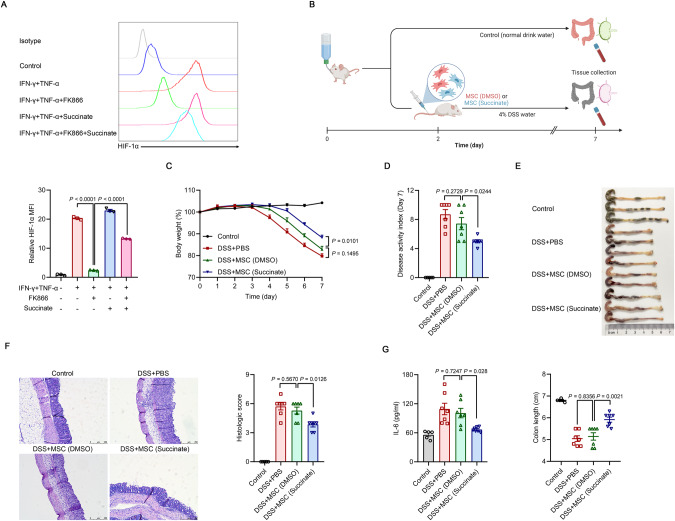
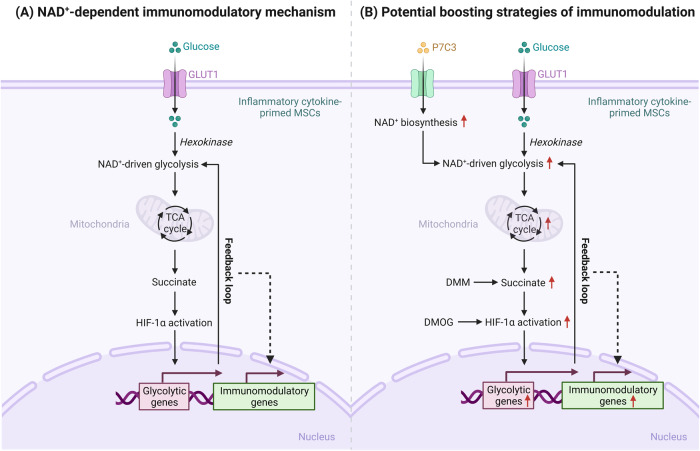
Mesenchymal stem/stromal cells (MSCs) possess robust immunoregulatory functions and are promising therapeutics for inflammatory disorders. This capacity is not innate but is activated or 'licensed' by inflammatory cytokines. The licensing mechanism remains unclear. Here, we examined whether inflammatory cytokines metabolically reprogrammed MSCs to confer this immunoregulatory capacity. In response to stimulation by inflammatory cytokines, MSCs exhibited a dramatic increase in the consumption of glucose, which was accompanied by an enhanced use of nicotinamide adenine dinucleotide (NAD+) and increased expression of nicotinamide phosphoribosyltransferase (NAMPT), a central enzyme in the salvage pathway for NAD+ production. When NAD+ synthesis was blocked by inhibiting or depleting NAMPT, the immunosuppressive function of MSCs induced by inflammatory cytokines was greatly attenuated. Consequently, when NAD+ metabolism in MSCs was perturbed, their therapeutic benefit was decreased in mice suffering from inflammatory bowel disease and acute liver injury. Further analysis revealed that NAMPT-driven production of NAD+ was critical for the inflammatory cytokine-induced increase in glycolysis in MSCs. Furthermore, the increase in glycolysis led to succinate accumulation in the tricarboxylic acid cycle, which led to hypoxia-inducible factor 1α (HIF-1α) stabilization and subsequently increased the transcription of key glycolytic genes, thereby persistently maintaining glycolytic flux. This study demonstrated that unlike its proinflammatory role in immune cells, NAD+ metabolism governs the anti-inflammatory function of MSCs during inflammation.
Keywords: Glycolysis; HIF-1α; Immunomodulation; Mesenchymal stem/stromal cells; NAD+ metabolism; Succinate.
© 2023. The Author(s), under exclusive licence to CSI and USTC.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
