

Sexual dimorphism in the social behaviour of Cntnap2-null mice correlates with disrupted synaptic connectivity and increased microglial activity in the anterior cingulate cortex
- PMID: 37582968
- PMCID: PMC10427688
- DOI: 10.1038/s42003-023-05215-0
Sexual dimorphism in the social behaviour of Cntnap2-null mice correlates with disrupted synaptic connectivity and increased microglial activity in the anterior cingulate cortex
Abstract
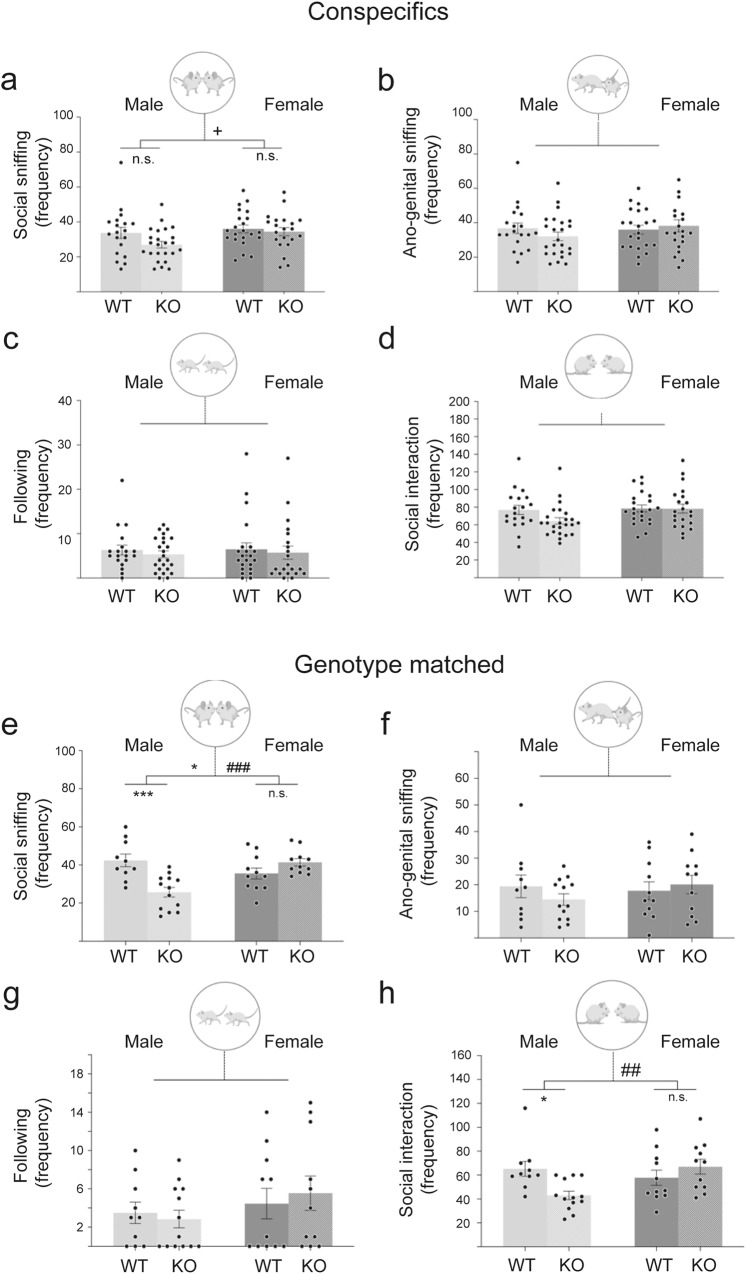
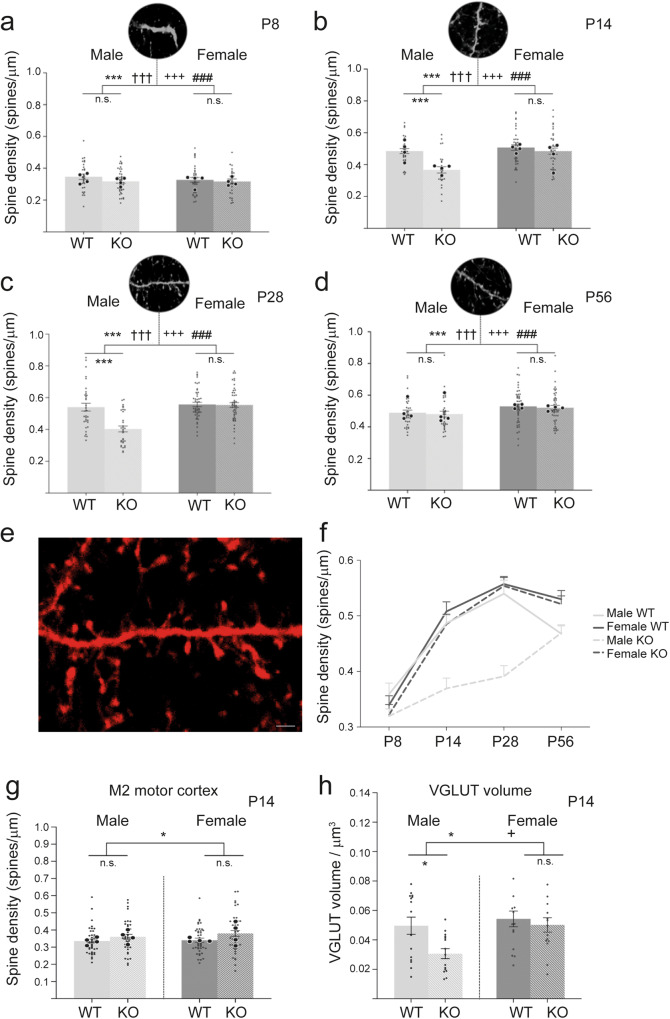
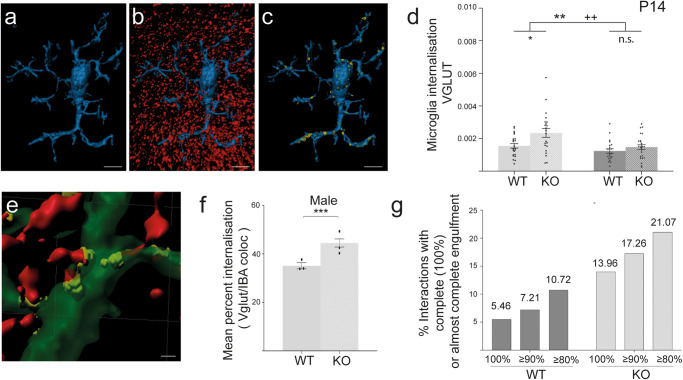
A biological understanding of the apparent sex bias in autism is lacking. Here we have identified Cntnap2 KO mice as a model system to help better understand this dimorphism. Using this model, we observed social deficits in juvenile male KO mice only. These male-specific social deficits correlated with reduced spine densities of Layer 2/3 and Layer 5 pyramidal neurons in the Anterior Cingulate Cortex, a forebrain region prominently associated with the control of social behaviour. Furthermore, in male KO mice, microglia showed an increased activated morphology and phagocytosis of synaptic structures compared to WT mice, whereas no differences were seen in female KO and WT mice. Our data suggest that sexually dimorphic microglial activity may be involved in the aetiology of ASD, disrupting the development of neural circuits that control social behaviour by overpruning synapses at a developmentally critical period.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Battle DE. Diagnostic and statistical manual of mental disorders (DSM) Codas. 2013;25:191–192. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
