

Metabolic reprogramming in inflammatory microglia indicates a potential way of targeting inflammation in Alzheimer's disease
- PMID: 37586250
- PMCID: PMC10457454
- DOI: 10.1016/j.redox.2023.102846
Metabolic reprogramming in inflammatory microglia indicates a potential way of targeting inflammation in Alzheimer's disease
Abstract
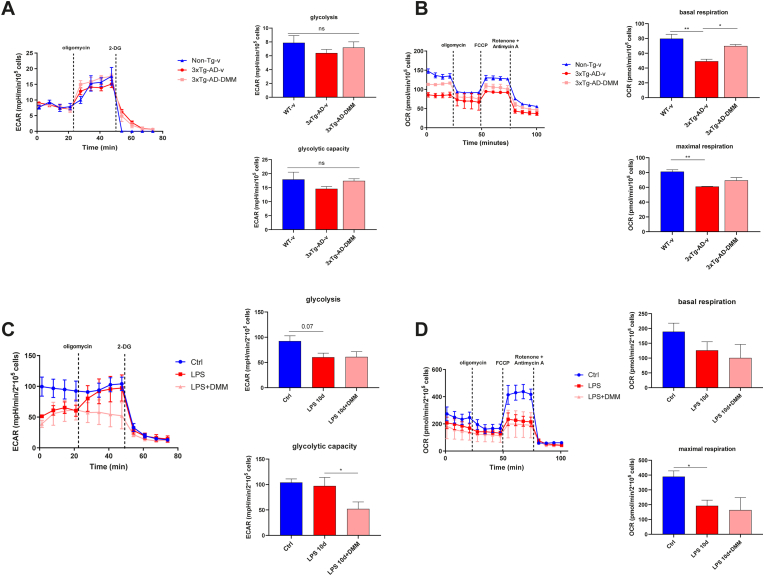
Microglia activation drives the pro-inflammatory activity in the early stages of Alzheimer's disease (AD). However, the mechanistic basis is elusive, and the hypothesis of targeting microglia to prevent AD onset is little explored. Here, we demonstrated that upon LPS exposure, microglia shift towards an energetic phenotype characterised by high glycolysis and high mitochondrial respiration with dysfunction. Although the activity of electron transport chain (ETC) complexes is boosted by LPS, this is mostly devoted to the generation of reactive oxygen species. We showed that by inhibiting succinate dehydrogenase (SDH) with dimethyl malonate (DMM), it is possible to modulate the LPS-induced metabolic rewiring, facilitating an anti-inflammatory phenotype. DMM improves mitochondrial function in a direct way and by reducing LPS-induced mitochondrial biogenesis. Moreover, the block of SDH with DMM inhibits the recruitment of hypoxia inducible-factor 1 α (HIF-1α), which mediates the induction of glycolysis and cytokine expression. Similar bioenergetic alterations were observed in the microglia isolated from AD mice (3xTg-AD), which present high levels of circulating LPS and brain toll-like receptor4 (TLR4). Moreover, this well-established model of AD was used to show a potential effect of SDH inhibition in vivo as DMM administration abrogated brain inflammation and modulated the microglia metabolic alterations of 3xTg-AD mice. The RNA-sequencing analysis from a public dataset confirmed the consistent transcription of genes encoding for ETC subunits in the microglia of AD mice (5xFAD). In conclusion, TLR4 activation promotes metabolic changes and the pro-inflammatory activity in microglia, and SDH might represent a promising therapeutic target to prevent AD development.
Keywords: 3xTg-AD mice; Alzheimer's disease; Bioenergetics; Dimethyl malonate; Immunometabolism; Macrophage; Microglia.
Copyright © 2023. Published by Elsevier B.V.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures







References
-
- Mucke L. Neuroscience: Alzheimer's disease. Nature. 2009;461:895–897. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
