

Platelet-derived exerkine CXCL4/platelet factor 4 rejuvenates hippocampal neurogenesis and restores cognitive function in aged mice
- PMID: 37587147
- PMCID: PMC10432533
- DOI: 10.1038/s41467-023-39873-9
Platelet-derived exerkine CXCL4/platelet factor 4 rejuvenates hippocampal neurogenesis and restores cognitive function in aged mice
Abstract
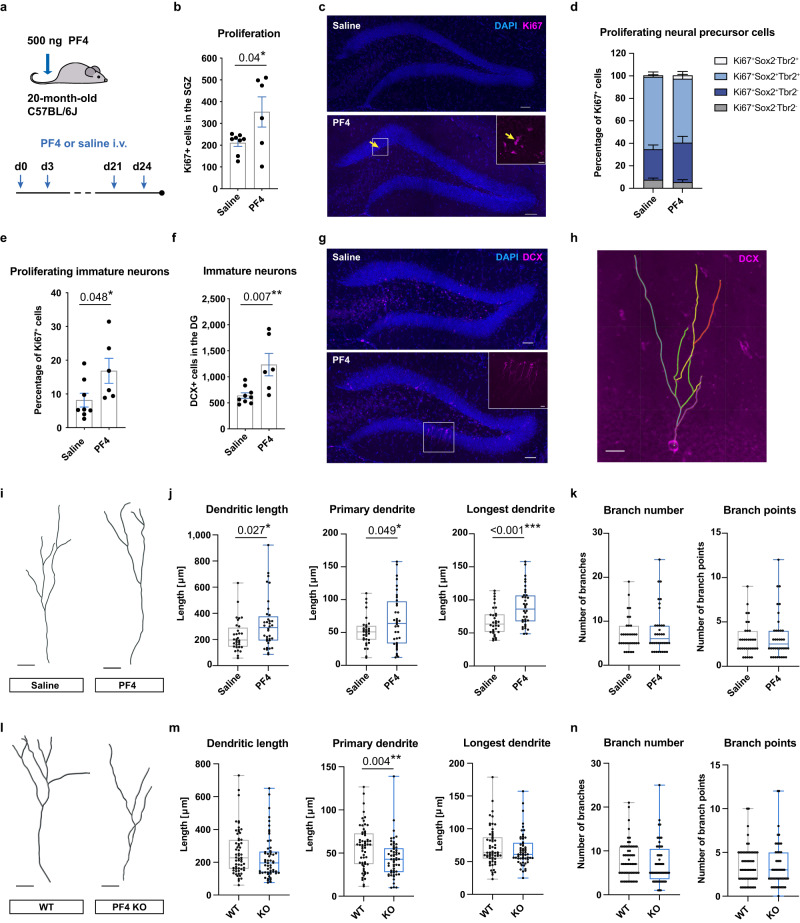
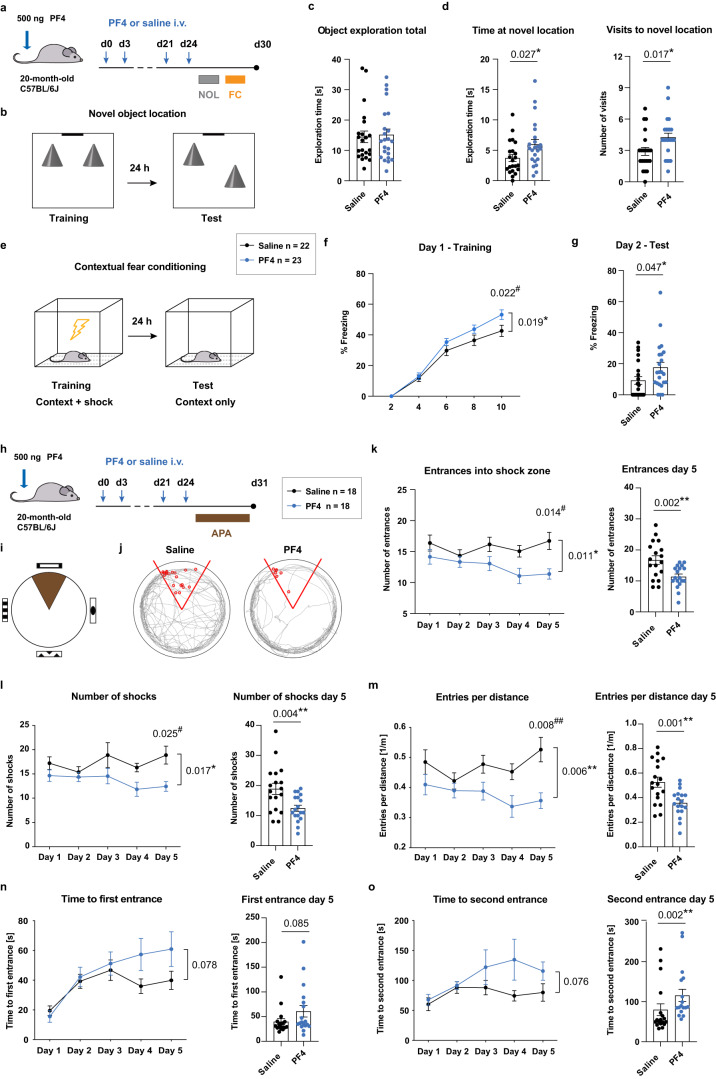
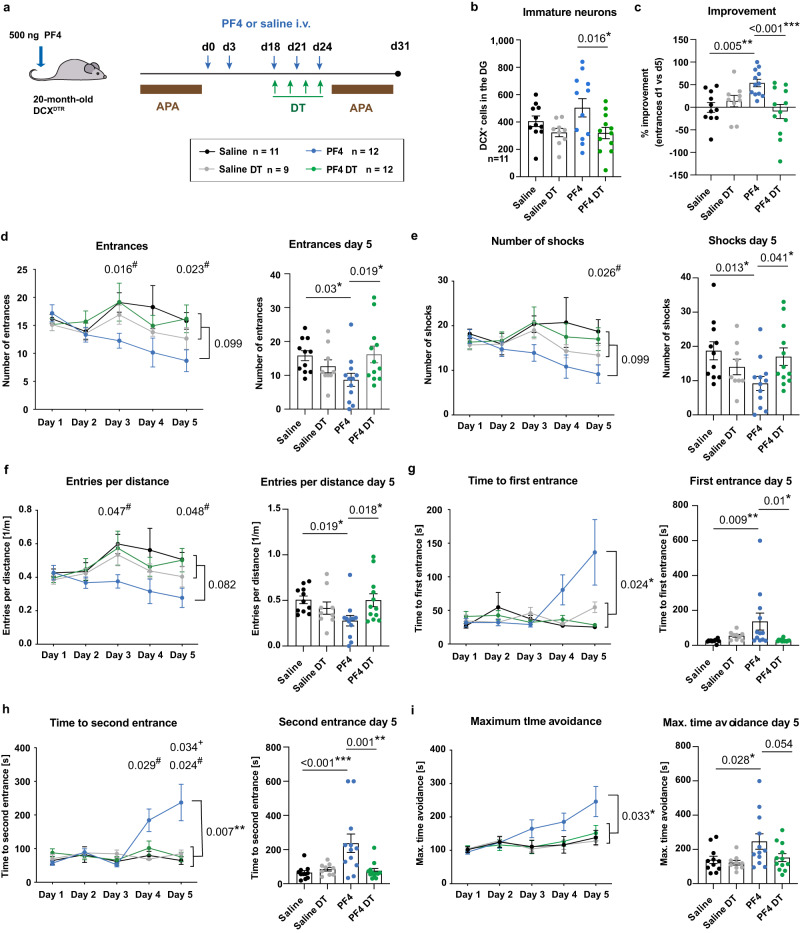
The beneficial effects of physical activity on brain ageing are well recognised, with exerkines, factors that are secreted into the circulation in response to exercise, emerging as likely mediators of this response. However, the source and identity of these exerkines remain unclear. Here we provide evidence that an anti-geronic exerkine is secreted by platelets. We show that platelets are activated by exercise and are required for the exercise-induced increase in hippocampal precursor cell proliferation in aged mice. We also demonstrate that increasing the systemic levels of the platelet-derived exerkine CXCL4/platelet factor 4 (PF4) ameliorates age-related regenerative and cognitive impairments in a hippocampal neurogenesis-dependent manner. Together these findings highlight the role of platelets in mediating the rejuvenating effects of exercise during physiological brain ageing.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
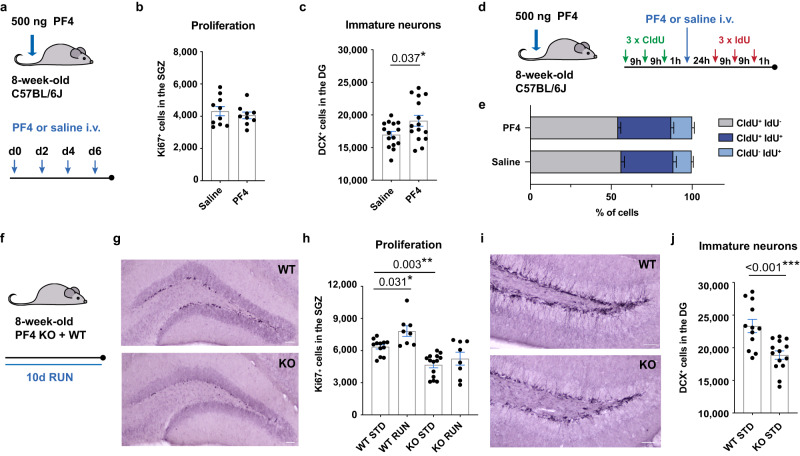
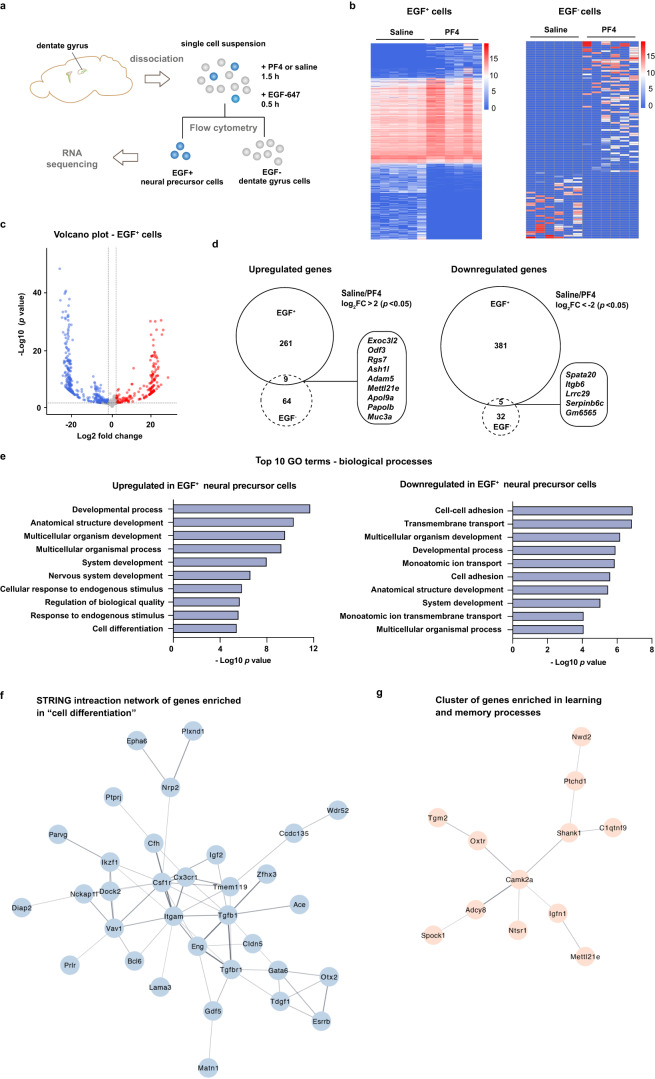
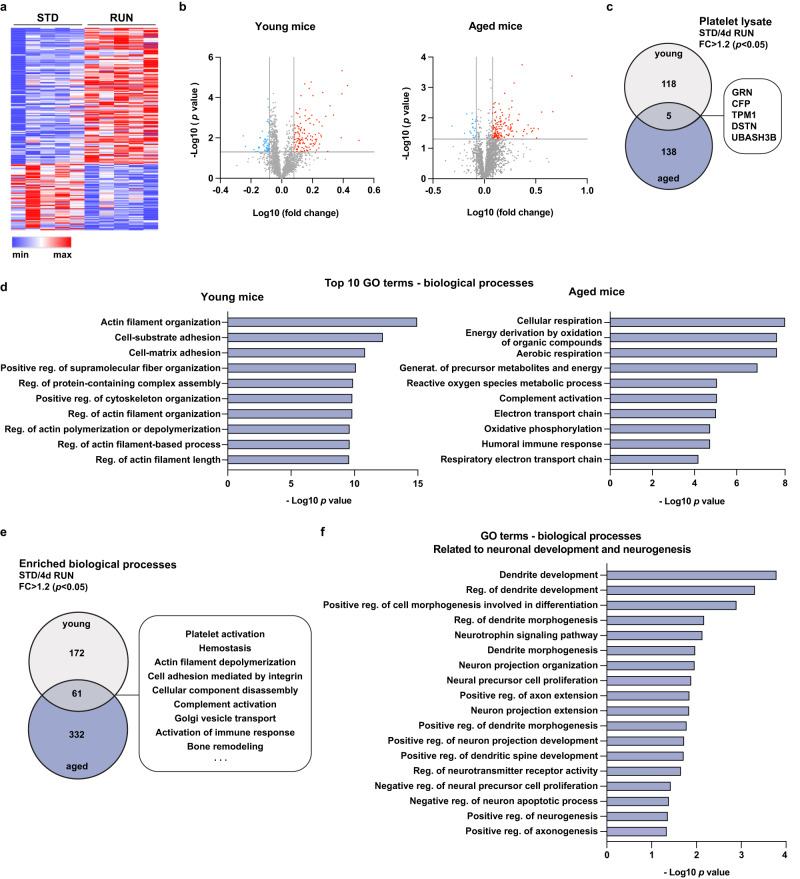
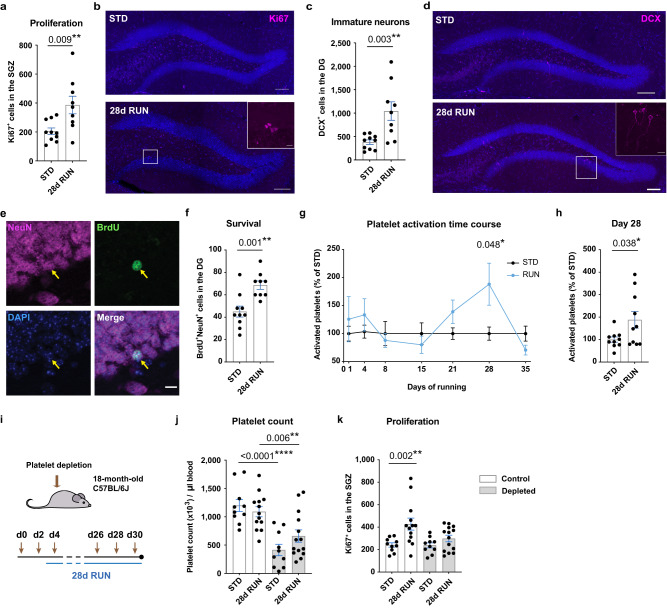
Comment in
-
Platelets rejuvenate the aging brain.Nat Cardiovasc Res. 2023 Oct;2(10):859. doi: 10.1038/s44161-023-00355-2. Nat Cardiovasc Res. 2023. PMID: 39196256 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
