

Mapping the T cell repertoire to a complex gut bacterial community
- PMID: 37587342
- PMCID: PMC10948025
- DOI: 10.1038/s41586-023-06431-8
Mapping the T cell repertoire to a complex gut bacterial community
Abstract
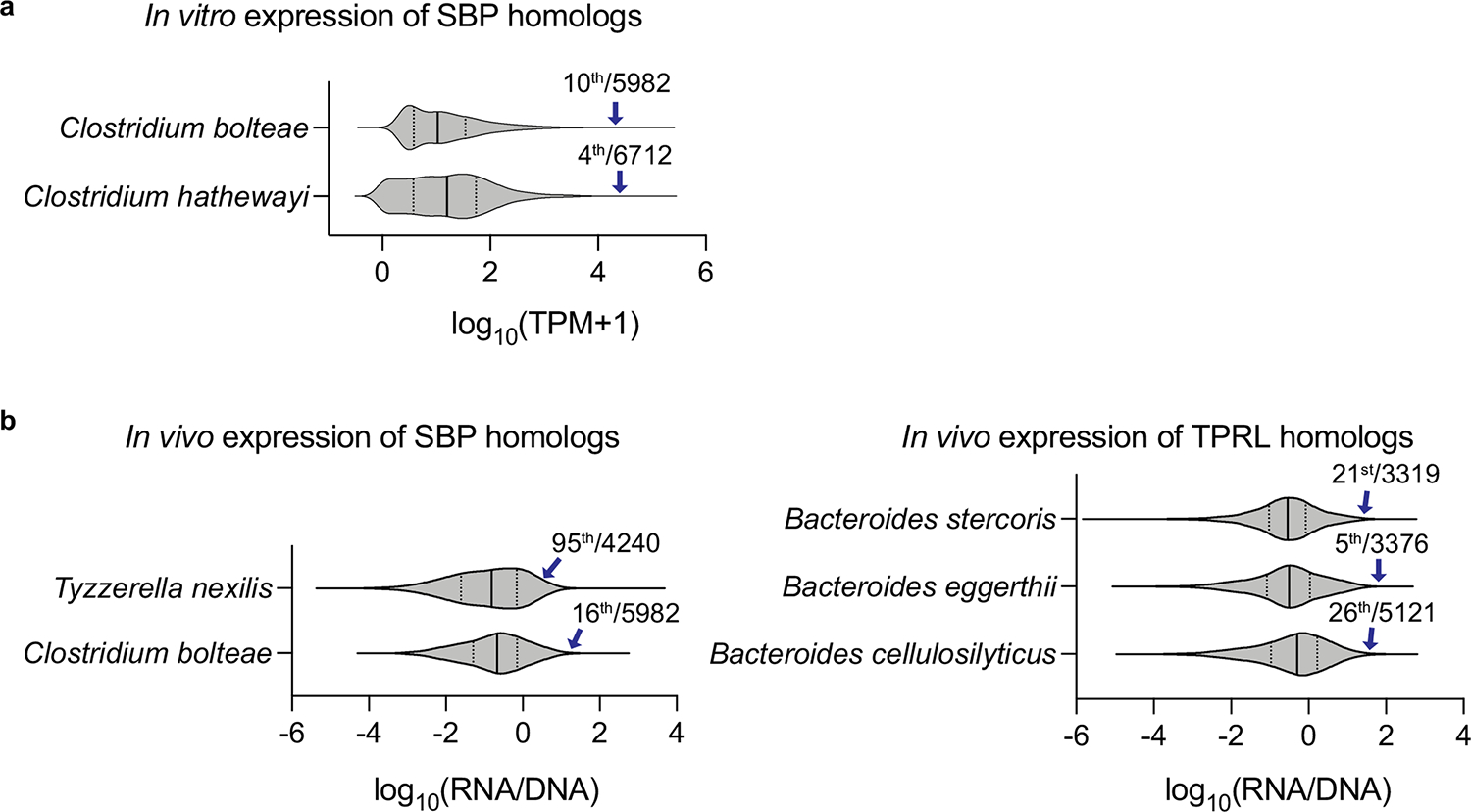
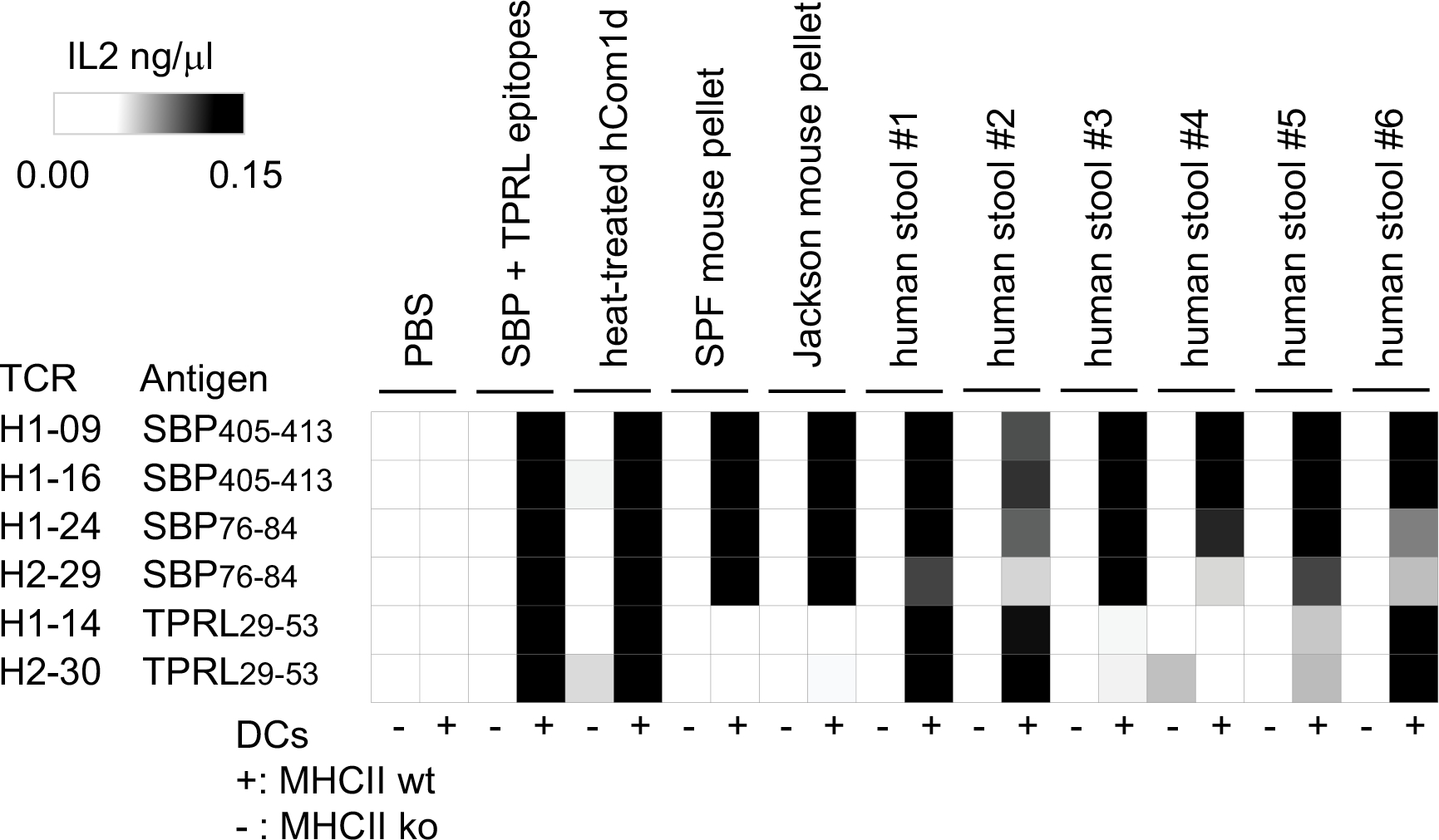
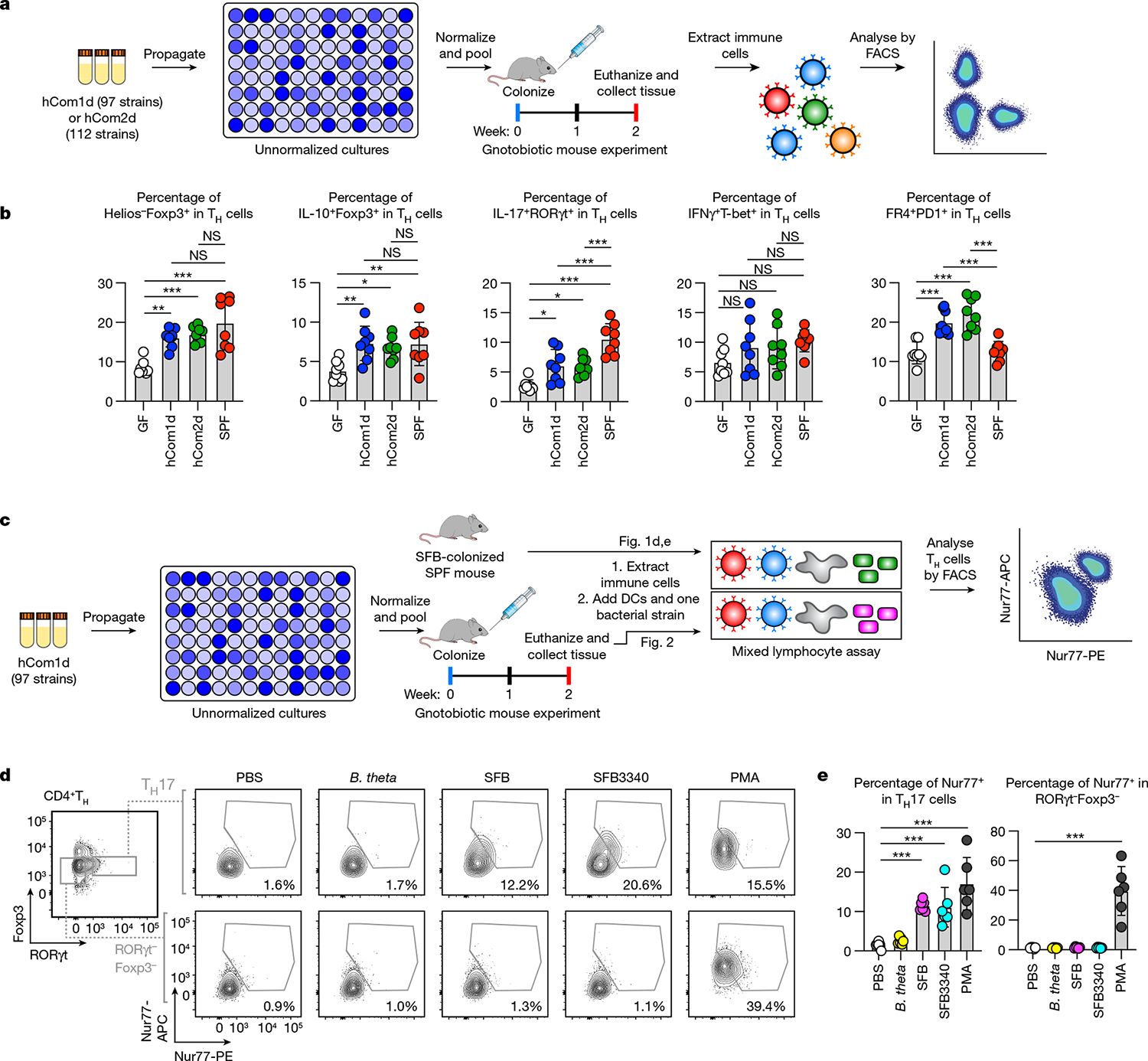
Certain bacterial strains from the microbiome induce a potent, antigen-specific T cell response1-5. However, the specificity of microbiome-induced T cells has not been explored at the strain level across the gut community. Here, we colonize germ-free mice with complex defined communities (roughly 100 bacterial strains) and profile T cell responses to each strain. The pattern of responses suggests that many T cells in the gut repertoire recognize several bacterial strains from the community. We constructed T cell hybridomas from 92 T cell receptor (TCR) clonotypes; by screening every strain in the community against each hybridoma, we find that nearly all the bacteria-specific TCRs show a one-to-many TCR-to-strain relationship, including 13 abundant TCR clonotypes that each recognize 18 Firmicutes. By screening three pooled bacterial genomic libraries, we discover that these 13 clonotypes share a single target: a conserved substrate-binding protein from an ATP-binding cassette transport system. Peripheral regulatory T cells and T helper 17 cells specific for an epitope from this protein are abundant in community-colonized and specific pathogen-free mice. Our work reveals that T cell recognition of commensals is focused on widely conserved, highly expressed cell-surface antigens, opening the door to new therapeutic strategies in which colonist-specific immune responses are rationally altered or redirected.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Figures













References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
