

Canonical and noncanonical Wnt signaling: Multilayered mediators, signaling mechanisms and major signaling crosstalk
- PMID: 37588235
- PMCID: PMC10425814
- DOI: 10.1016/j.gendis.2023.01.030
Canonical and noncanonical Wnt signaling: Multilayered mediators, signaling mechanisms and major signaling crosstalk
Abstract
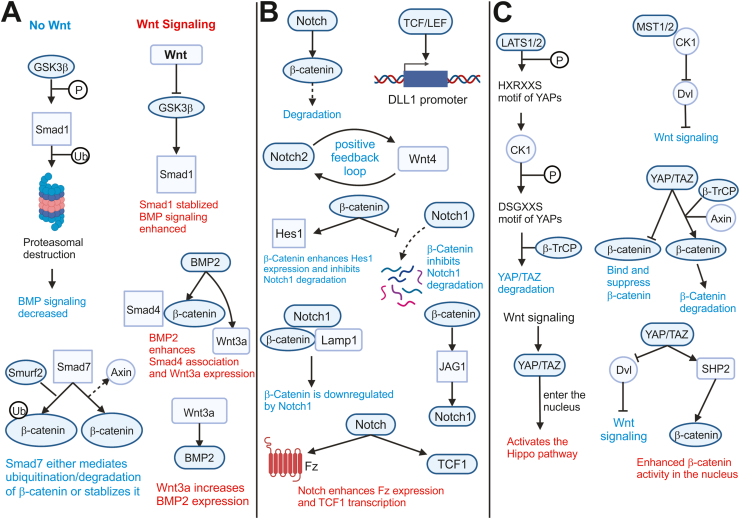
Wnt signaling plays a major role in regulating cell proliferation and differentiation. The Wnt ligands are a family of 19 secreted glycoproteins that mediate their signaling effects via binding to Frizzled receptors and LRP5/6 coreceptors and transducing the signal either through β-catenin in the canonical pathway or through a series of other proteins in the noncanonical pathway. Many of the individual components of both canonical and noncanonical Wnt signaling have additional functions throughout the body, establishing the complex interplay between Wnt signaling and other signaling pathways. This crosstalk between Wnt signaling and other pathways gives Wnt signaling a vital role in many cellular and organ processes. Dysregulation of this system has been implicated in many diseases affecting a wide array of organ systems, including cancer and embryological defects, and can even cause embryonic lethality. The complexity of this system and its interacting proteins have made Wnt signaling a target for many therapeutic treatments. However, both stimulatory and inhibitory treatments come with potential risks that need to be addressed. This review synthesized much of the current knowledge on the Wnt signaling pathway, beginning with the history of Wnt signaling. It thoroughly described the different variants of Wnt signaling, including canonical, noncanonical Wnt/PCP, and the noncanonical Wnt/Ca2+ pathway. Further description involved each of its components and their involvement in other cellular processes. Finally, this review explained the various other pathways and processes that crosstalk with Wnt signaling.
Keywords: Canonical Wnt; Noncanonical Wnt; Signal transduction; Signaling crosstalk; β-catenin.
© 2023 The Authors. Publishing services by Elsevier B.V. on behalf of KeAi Communications Co., Ltd.
Figures




References
-
- Nusse R., Varmus H.E. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell. 1982;31(1):99–109. - PubMed
-
- Sharma R.P., Chopra V.L. Effect of the Wingless (wg1) mutation on the wing and haltere development in Drosophila melanogaster. Dev Biol. 1976;48(2):461–465. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
