

An interaction between OTULIN and SCRIB uncovers roles for linear ubiquitination in planar cell polarity
- PMID: 37589075
- PMCID: PMC10445738
- DOI: 10.1242/dmm.049762
An interaction between OTULIN and SCRIB uncovers roles for linear ubiquitination in planar cell polarity
Abstract
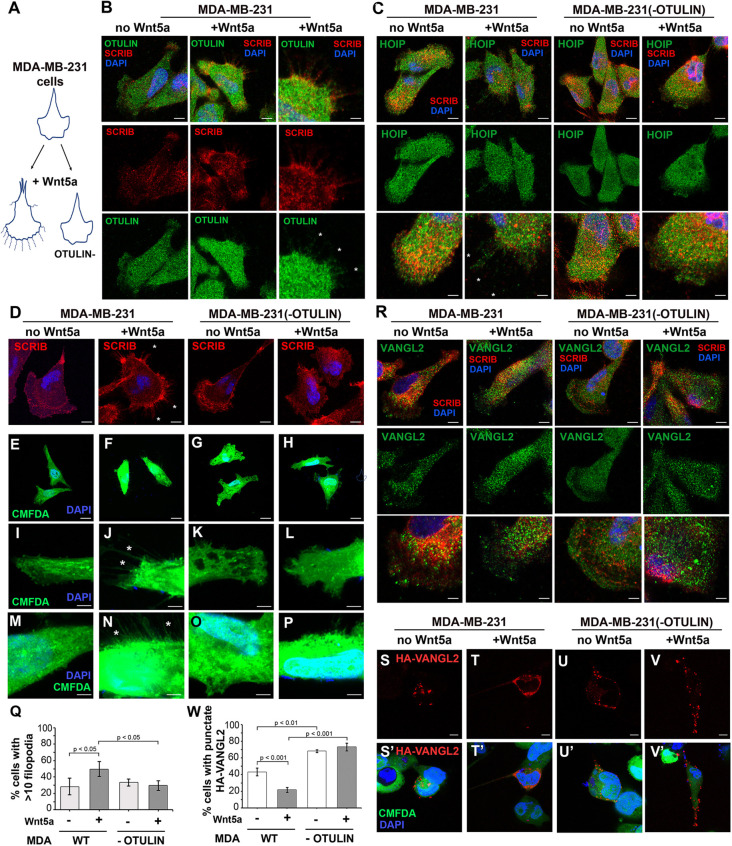
Planar cell polarity (PCP) plays critical roles in developmental and homeostatic processes. Membrane presentation of PCP complexes containing Van Gogh-like (VANGL) transmembrane proteins is central to PCP and can be directed by the scaffold protein scribble (SCRIB). The role atypical linear ubiquitin (Met1-Ub) chains might play in PCP is unknown. Here, HEK293 cell-based interactomic analyses of the Met1-Ub deubiquitinase OTULIN revealed that OTULIN can interact with SCRIB. Moreover, Met1-Ub chains associated with VANGL2 and PRICKLE1, but not SCRIB, can direct VANGL2 surface presentation. Mouse embryos lacking Otulin showed variable neural tube malformations, including rare open neural tubes, a deficit associated with PCP disruption in mice. In Madin-Darby canine kidney cells, in which the enrichment of VANGL2-GFP proteins at cell-cell contacts represents activated PCP complexes, endogenous OTULIN was recruited to these sites. In the human MDA-MB-231 breast cancer cell model, OTULIN loss caused deficits in Wnt5a-induced filopodia extension and trafficking of transfected HA-VANGL2. Taken together, these findings support a role for linear (de)ubiquitination in PCP signaling. The association of Met1-Ub chains with PCP complex components offers new opportunities for integrating PCP signaling with OTULIN-dependent immune and inflammatory pathways.
Keywords: Linear ubiquitin; Linear ubiquitin assembly complex; Mouse genetics; OTULIN; Planar cell polarity; Scribble.
© 2023. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures





References
-
- Bassuk, A. G., Wallace, R. H., Buhr, A., Buller, A. R., Afawi, Z., Shimojo, M., Miyata, S., Chen, S., Gonzalez-Alegre, P., Griesbach, H. L.et al. (2008). A homozygous mutation in human PRICKLE1 causes an autosomal-recessive progressive myoclonus epilepsy-ataxia syndrome. Am. J. Hum. Genet. 83, 572-581. 10.1016/j.ajhg.2008.10.003 - DOI - PMC - PubMed
-
- Belotti, E., Polanowska, J., Daulat, A. M., Audebert, S., Thome, V., Lissitzky, J. C., Lembo, F., Blibek, K., Omi, S., Lenfant, N.et al. (2013). The human PDZome: a gateway to PSD95-Disc large-zonula occludens (PDZ)-mediated functions. Mol. Cell. Proteomics 12, 2587-2603. 10.1074/mcp.O112.021022 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
