

Yolk sac cell atlas reveals multiorgan functions during human early development
- PMID: 37590359
- PMCID: PMC7614978
- DOI: 10.1126/science.add7564
Yolk sac cell atlas reveals multiorgan functions during human early development
Abstract
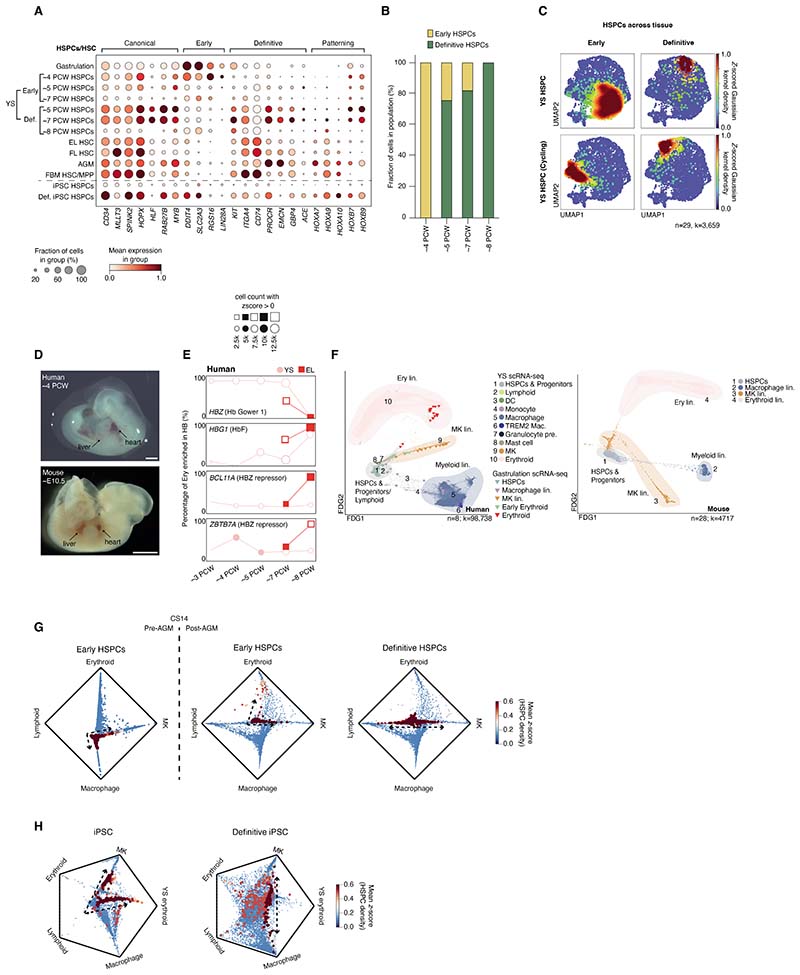
The extraembryonic yolk sac (YS) ensures delivery of nutritional support and oxygen to the developing embryo but remains ill-defined in humans. We therefore assembled a comprehensive multiomic reference of the human YS from 3 to 8 postconception weeks by integrating single-cell protein and gene expression data. Beyond its recognized role as a site of hematopoiesis, we highlight roles in metabolism, coagulation, vascular development, and hematopoietic regulation. We reconstructed the emergence and decline of YS hematopoietic stem and progenitor cells from hemogenic endothelium and revealed a YS-specific accelerated route to macrophage production that seeds developing organs. The multiorgan functions of the YS are superseded as intraembryonic organs develop, effecting a multifaceted relay of vital functions as pregnancy proceeds.
Conflict of interest statement
Figures





Comment in
-
Illuminating the human yolk sac through single-cell omics.Nat Rev Immunol. 2023 Nov;23(11):701. doi: 10.1038/s41577-023-00957-8. Nat Rev Immunol. 2023. PMID: 37813930 No abstract available.
References
-
- Palis J, Malik J, Mcgrath KE, Kingsley PD. Primitive erythropoiesis in the mammalian embryo. The International Journal of Developmental Biology. 2010;54:1011–1018. - PubMed
MeSH terms
Grants and funding
- MC_PC_17230/MRC_/Medical Research Council/United Kingdom
- 215116/WT_/Wellcome Trust/United Kingdom
- 206328/WT_/Wellcome Trust/United Kingdom
- 206328/Z/17/Z/WT_/Wellcome Trust/United Kingdom
- MC_UU_00029/5/MRC_/Medical Research Council/United Kingdom
- MR/S036113/1/MRC_/Medical Research Council/United Kingdom
- 206194/WT_/Wellcome Trust/United Kingdom
- R01 HL151777/HL/NHLBI NIH HHS/United States
- MR/M008975/1/MRC_/Medical Research Council/United Kingdom
- MR/R006237/1/MRC_/Medical Research Council/United Kingdom
- 107931/WT_/Wellcome Trust/United Kingdom
- 211276/WT_/Wellcome Trust/United Kingdom
- 107630/WT_/Wellcome Trust/United Kingdom
- 220540/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
