

In silico protein interaction screening uncovers DONSON's role in replication initiation
- PMID: 37590370
- PMCID: PMC10801813
- DOI: 10.1126/science.adi3448
In silico protein interaction screening uncovers DONSON's role in replication initiation
Abstract
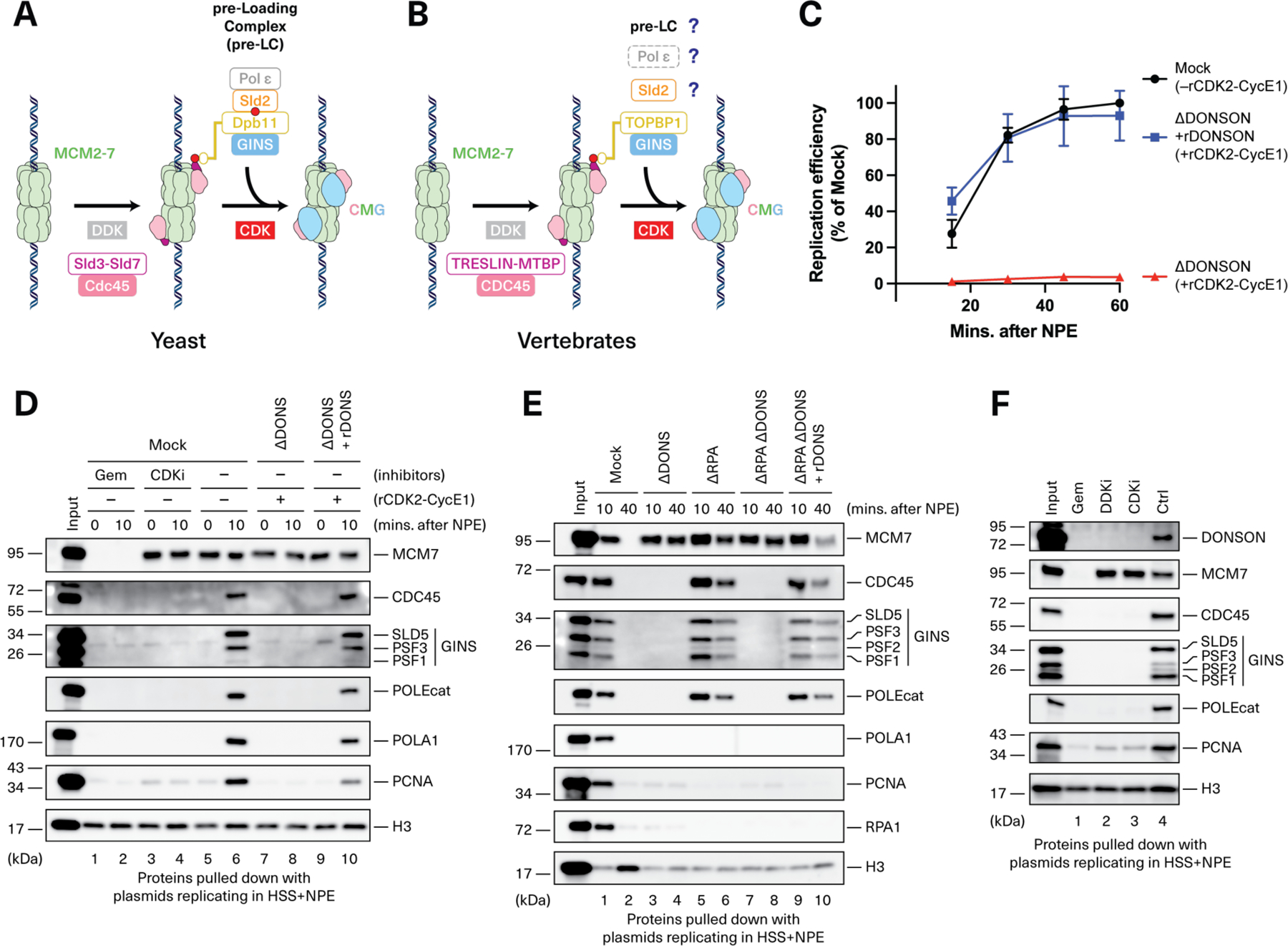
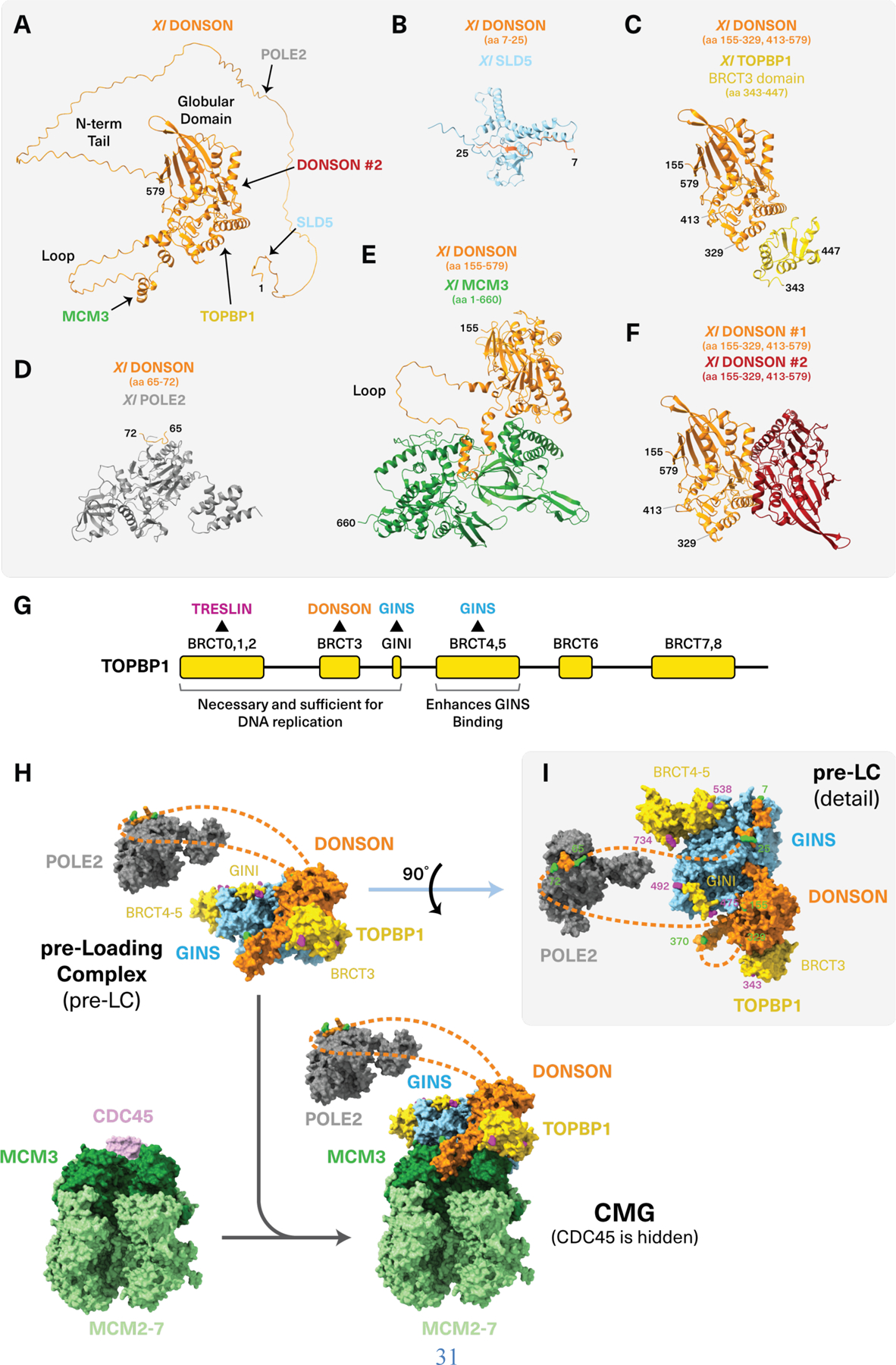
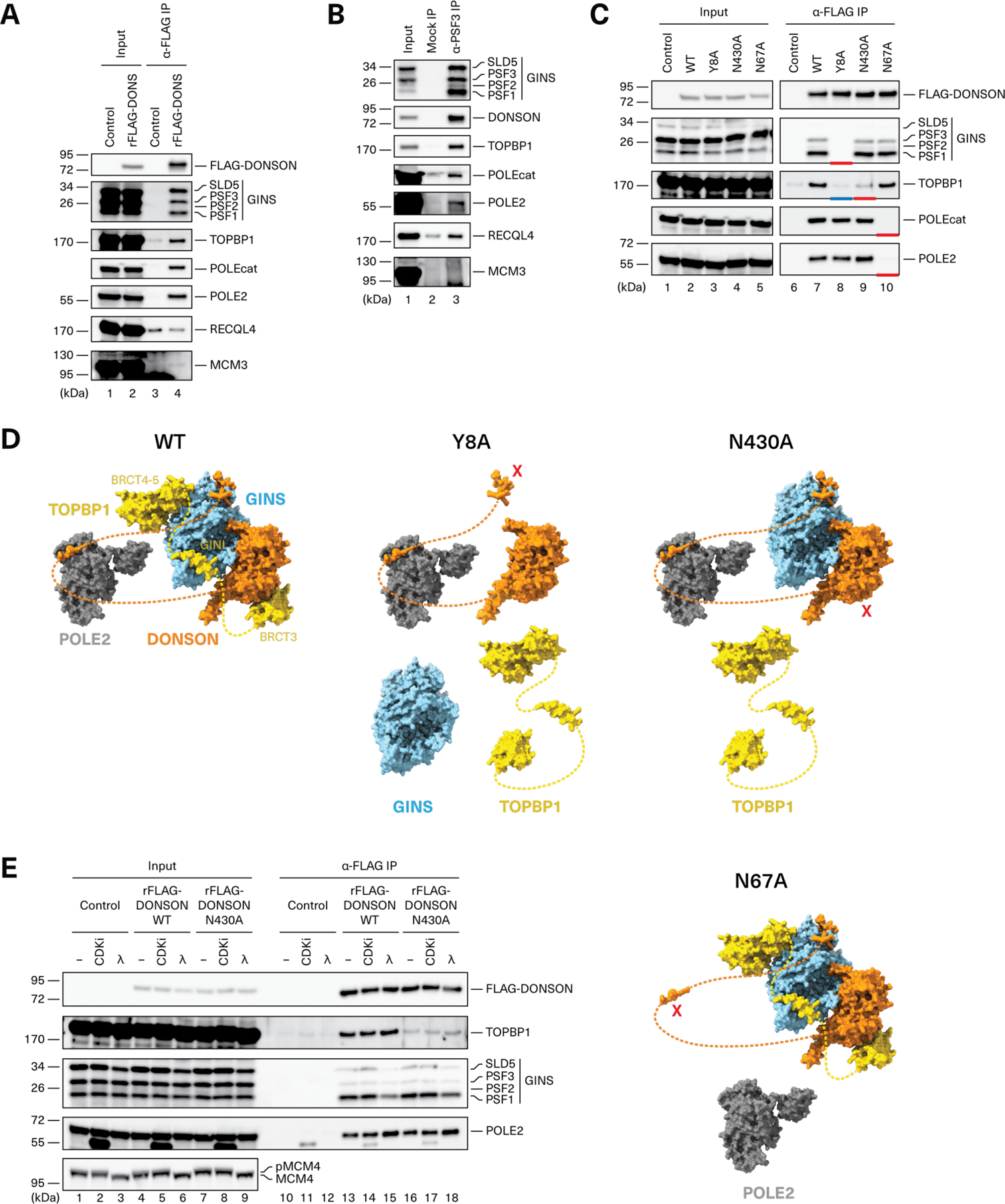
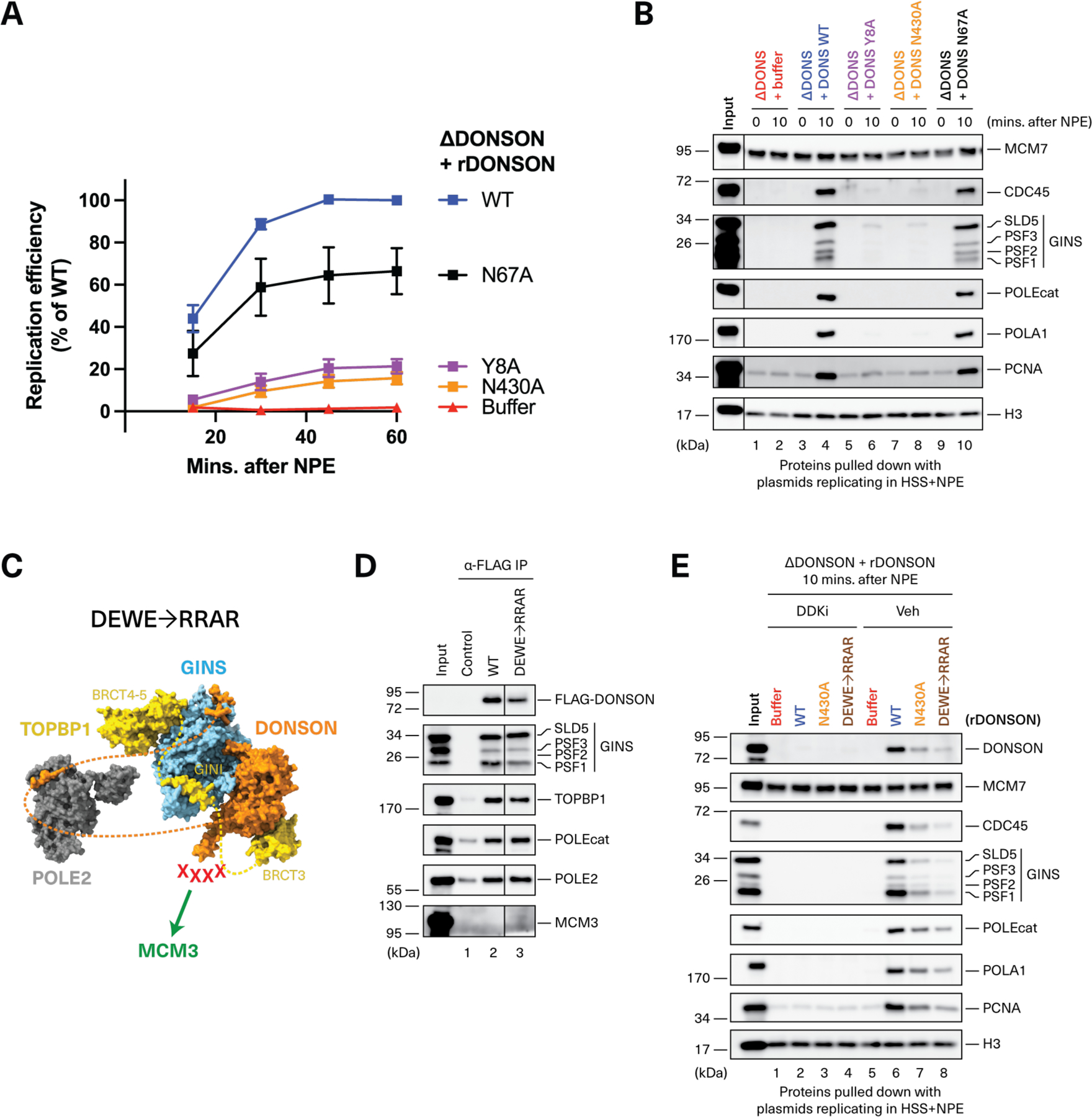
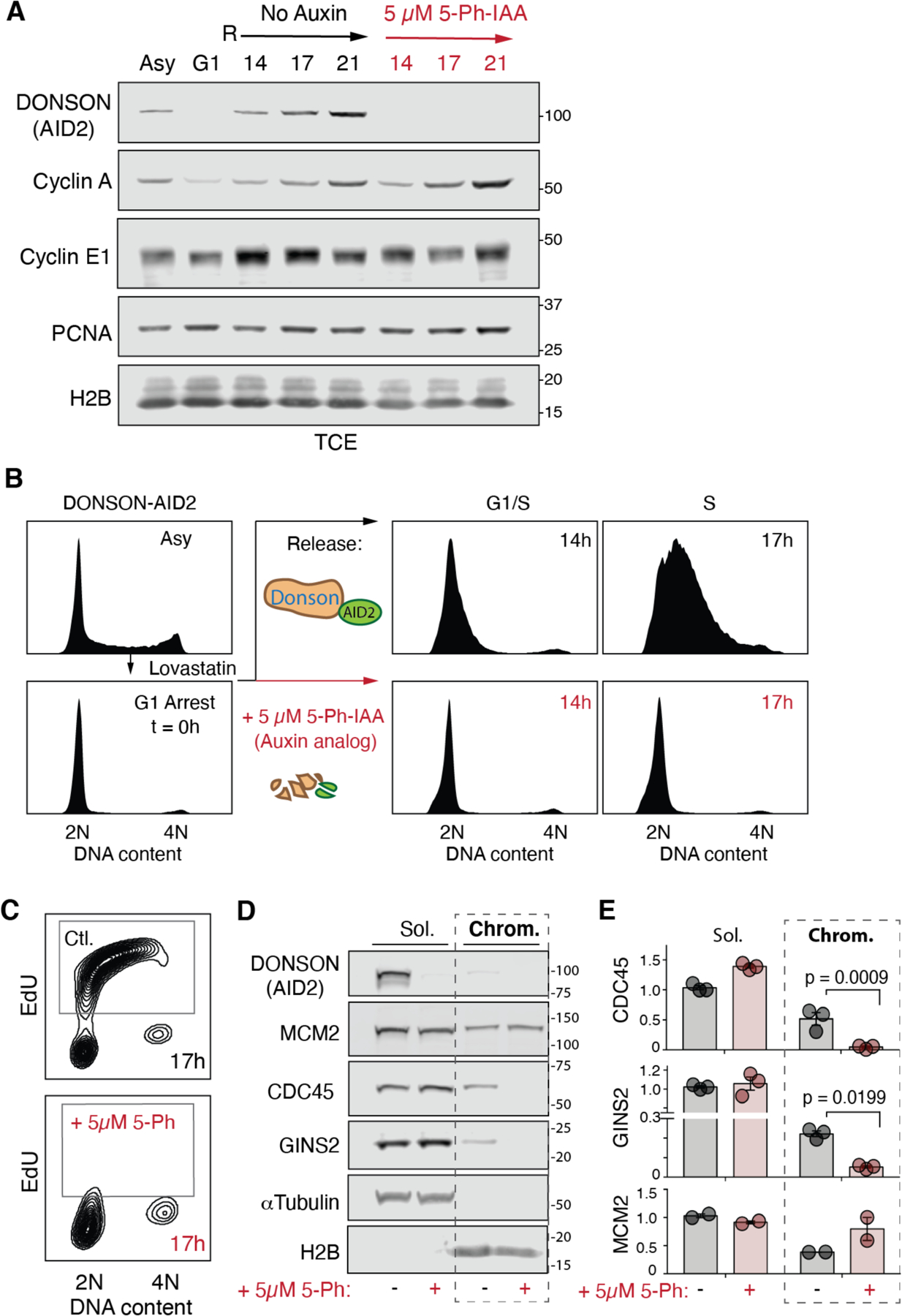
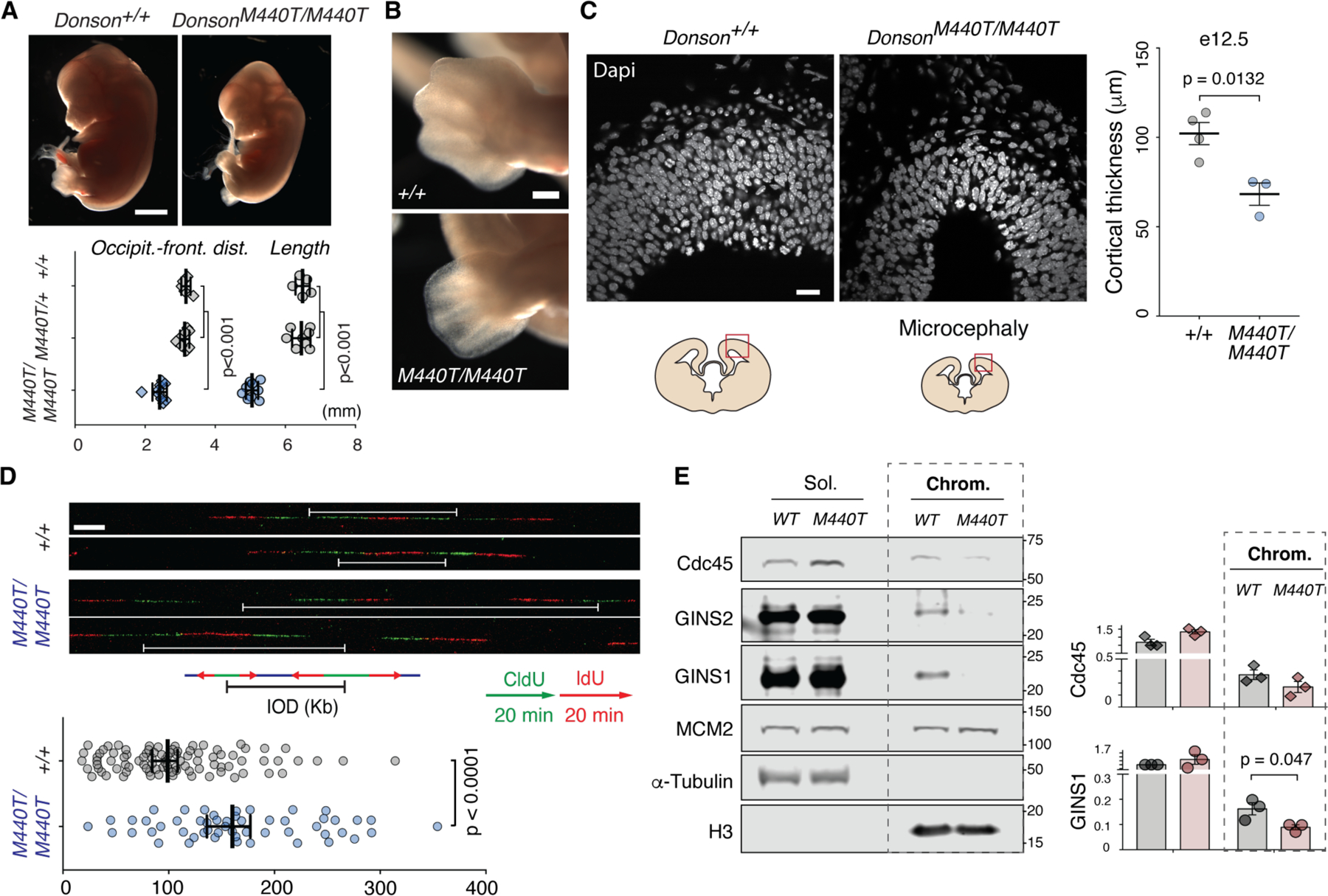
CDC45-MCM2-7-GINS (CMG) helicase assembly is the central event in eukaryotic replication initiation. In yeast, a multi-subunit "pre-loading complex" (pre-LC) accompanies GINS to chromatin-bound MCM2-7, leading to CMG formation. Here, we report that DONSON, a metazoan protein mutated in microcephalic primordial dwarfism, is required for CMG assembly in vertebrates. Using AlphaFold to screen for protein-protein interactions followed by experimental validation, we show that DONSON scaffolds a vertebrate pre-LC containing GINS, TOPBP1, and DNA pol ε. Our evidence suggests that DONSON docks the pre-LC onto MCM2-7, delivering GINS to its binding site in CMG. A patient-derived DONSON mutation compromises CMG assembly and recapitulates microcephalic dwarfism in mice. These results unify our understanding of eukaryotic replication initiation, implicate defective CMG assembly in microcephalic dwarfism, and illustrate how in silico protein-protein interaction screening accelerates mechanistic discovery.
Figures
References
-
- Costa A, Diffley JFX, The Initiation of Eukaryotic DNA Replication. Annu Rev Biochem 91, 107–131 (2022). - PubMed
-
- Tanaka S et al. CDK-dependent phosphorylation of Sld2 and Sld3 initiates DNA replication in budding yeast. Nature 445, 328–332 (2007). - PubMed
-
- Zegerman P, Diffley JF, Phosphorylation of Sld2 and Sld3 by cyclin-dependent kinases promotes DNA replication in budding yeast. Nature 445, 281–285 (2007). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
