

Cell-specific and shared regulatory elements control a multigene locus active in mammary and salivary glands
- PMID: 37591874
- PMCID: PMC10435465
- DOI: 10.1038/s41467-023-40712-0
Cell-specific and shared regulatory elements control a multigene locus active in mammary and salivary glands
Abstract
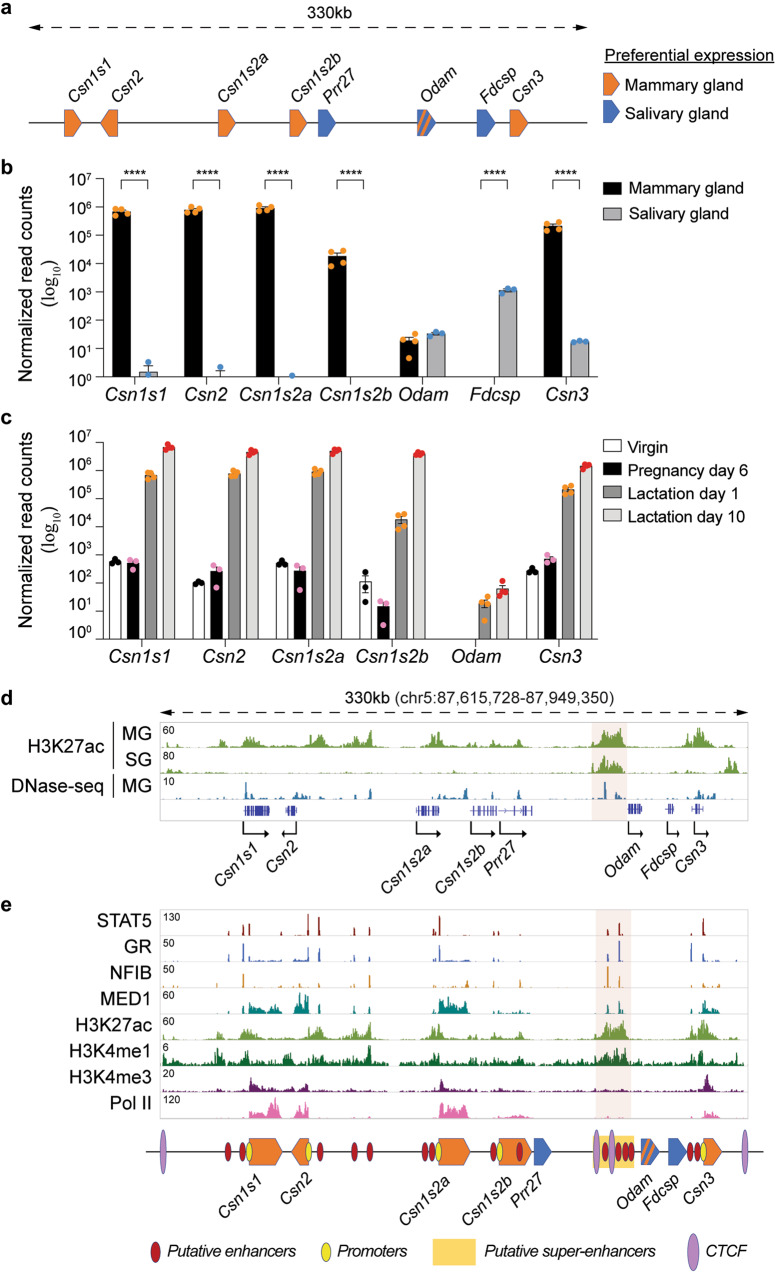
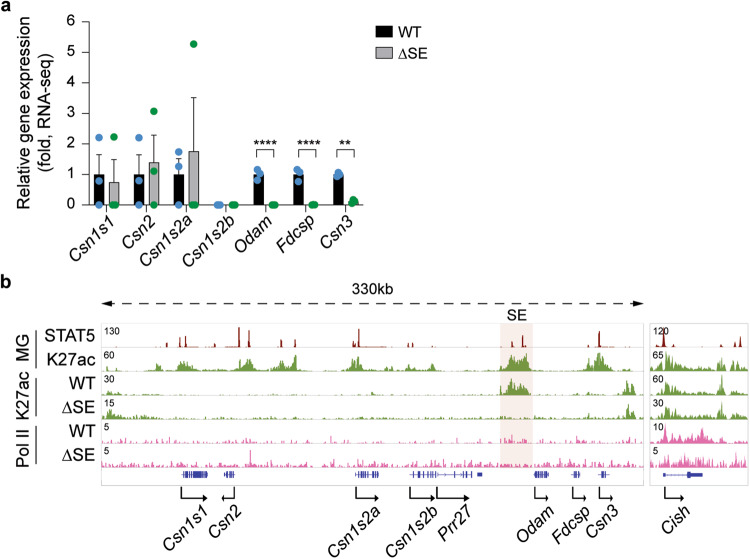
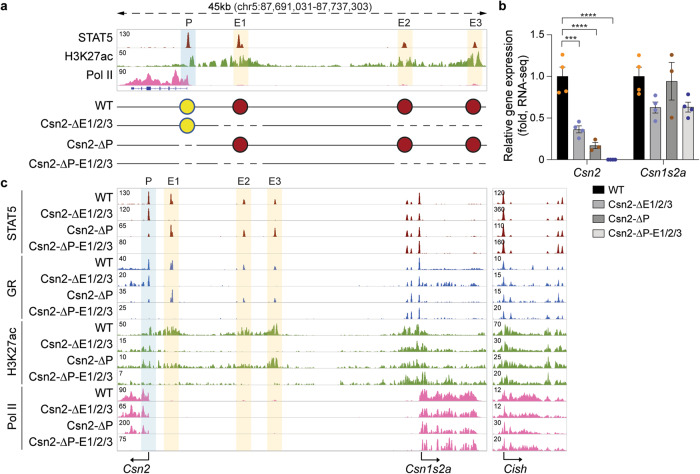
Regulation of high-density loci harboring genes with different cell-specificities remains a puzzle. Here we investigate a locus that evolved through gene duplication and contains eight genes and 20 candidate regulatory elements, including one super-enhancer. Casein genes (Csn1s1, Csn2, Csn1s2a, Csn1s2b, Csn3) are expressed in mammary glands, induced 10,000-fold during pregnancy and account for 50% of mRNAs during lactation, Prr27 and Fdcsp are salivary-specific and Odam has dual specificity. We probed the function of 12 candidate regulatory elements, individually and in combination, in the mouse genome. The super-enhancer is essential for the expression of Csn3, Csn1s2b, Odam and Fdcsp but largely dispensable for Csn1s1, Csn2 and Csn1s2a. Csn3 activation also requires its own local enhancer. Synergism between local enhancers and cytokine-responsive promoter elements facilitates activation of Csn2 during pregnancy. Our work identifies the regulatory complexity of a multigene locus with an ancestral super-enhancer active in mammary and salivary tissue and local enhancers and promoter elements unique to mammary tissue.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures






Update of
-
Cell-specific and shared enhancers control a high-density multi-gene locus active in mammary and salivary glands.Res Sq [Preprint]. 2023 Feb 7:rs.3.rs-2533579. doi: 10.21203/rs.3.rs-2533579/v1. Res Sq. 2023. Update in: Nat Commun. 2023 Aug 17;14(1):4992. doi: 10.1038/s41467-023-40712-0. PMID: 36789414 Free PMC article. Updated. Preprint.
-
Cell-specific and shared enhancers control a high-density multi-gene locus active in mammary and salivary glands.bioRxiv [Preprint]. 2023 Feb 7:2023.02.06.527373. doi: 10.1101/2023.02.06.527373. bioRxiv. 2023. Update in: Nat Commun. 2023 Aug 17;14(1):4992. doi: 10.1038/s41467-023-40712-0. PMID: 36945503 Free PMC article. Updated. Preprint.
Similar articles
-
Cell-specific and shared enhancers control a high-density multi-gene locus active in mammary and salivary glands.bioRxiv [Preprint]. 2023 Feb 7:2023.02.06.527373. doi: 10.1101/2023.02.06.527373. bioRxiv. 2023. Update in: Nat Commun. 2023 Aug 17;14(1):4992. doi: 10.1038/s41467-023-40712-0. PMID: 36945503 Free PMC article. Updated. Preprint.
-
Cell-specific and shared enhancers control a high-density multi-gene locus active in mammary and salivary glands.Res Sq [Preprint]. 2023 Feb 7:rs.3.rs-2533579. doi: 10.21203/rs.3.rs-2533579/v1. Res Sq. 2023. Update in: Nat Commun. 2023 Aug 17;14(1):4992. doi: 10.1038/s41467-023-40712-0. PMID: 36789414 Free PMC article. Updated. Preprint.
-
Redundant and non-redundant cytokine-activated enhancers control Csn1s2b expression in the lactating mouse mammary gland.Nat Commun. 2021 Apr 14;12(1):2239. doi: 10.1038/s41467-021-22500-w. Nat Commun. 2021. PMID: 33854063 Free PMC article.
-
STAT5-Driven Enhancers Tightly Control Temporal Expression of Mammary-Specific Genes.J Mammary Gland Biol Neoplasia. 2019 Mar;24(1):61-71. doi: 10.1007/s10911-018-9418-y. Epub 2018 Oct 17. J Mammary Gland Biol Neoplasia. 2019. PMID: 30328555 Review.
-
Multispecies comparison of the casein gene loci and evolution of casein gene family.J Mammary Gland Biol Neoplasia. 2002 Jul;7(3):327-45. doi: 10.1023/a:1022808918013. J Mammary Gland Biol Neoplasia. 2002. PMID: 12751895 Review.
Cited by
-
Molecular Diversity of the Casein Gene Cluster in Bovidae: Insights from SNP Microarray Analysis.Animals (Basel). 2024 Oct 19;14(20):3034. doi: 10.3390/ani14203034. Animals (Basel). 2024. PMID: 39457964 Free PMC article.
-
Emergence of saliva protein genes in the secretory calcium-binding phosphoprotein (SCPP) locus and accelerated evolution in primates.bioRxiv [Preprint]. 2024 Feb 15:2024.02.14.580359. doi: 10.1101/2024.02.14.580359. bioRxiv. 2024. PMID: 38405690 Free PMC article. Preprint.
-
The Effect of CDKN1A on the Expression of Genes Related to Milk Protein and Milk Fat Synthesis in Bovine Mammary Epithelial Cells.Vet Sci. 2025 Jun 1;12(6):534. doi: 10.3390/vetsci12060534. Vet Sci. 2025. PMID: 40559771 Free PMC article.
-
STAT5B leukemic mutations, altering SH2 tyrosine 665, have opposing impacts on immune gene programs.Life Sci Alliance. 2025 Apr 14;8(7):e202503222. doi: 10.26508/lsa.202503222. Print 2025 Jul. Life Sci Alliance. 2025. PMID: 40228864 Free PMC article.
-
Autologous porcine VRAM flap model for VCA research.Front Transplant. 2024 Dec 6;3:1504959. doi: 10.3389/frtra.2024.1504959. eCollection 2024. Front Transplant. 2024. PMID: 39712036 Free PMC article.
References
-
- Oftedal OT. The mammary gland and its origin during synapsid evolution. J. Mammary Gland Biol. Neoplasia. 2002;7:225–252. - PubMed
-
- Kawasaki K, Buchanan AV, Weiss KM. Biomineralization in humans: making the hard choices in life. Annu. Rev. Genet. 2009;43:119–142. - PubMed
-
- Kawasaki K. The SCPP gene family and the complexity of hard tissues in vertebrates. Cells Tissues Organs. 2011;194:108–112. - PubMed
-
- Hennighausen LG, Sippel AE. Characterization and cloning of the mRNAs specific for the lactating mouse mammary gland. Eur. J. Biochem. 1982;125:131–141. - PubMed
-
- Rijnkels M, Elnitski L, Miller W, Rosen JM. Multispecies comparative analysis of a mammalian-specific genomic domain encoding secretory proteins. Genomics. 2003;82:417–432. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
