

ApoE expression in macrophages communicates immunometabolic signaling that controls hyperlipidemia-driven hematopoiesis & inflammation via extracellular vesicles
- PMID: 37593979
- PMCID: PMC10436255
- DOI: 10.1002/jev2.12345
ApoE expression in macrophages communicates immunometabolic signaling that controls hyperlipidemia-driven hematopoiesis & inflammation via extracellular vesicles
Erratum in
-
Correction to "ApoE expression in macrophages communicates immunometabolic signaling that controls hyperlipidemia-driven hematopoiesis & inflammation via extracellular vesicles".J Extracell Vesicles. 2023 Nov;12(11):e12375. doi: 10.1002/jev2.12375. J Extracell Vesicles. 2023. PMID: 37885049 Free PMC article. No abstract available.
-
Correction to article pagination in the Journal of Extracellular Vesicles.J Extracell Vesicles. 2024 May;13(5):e12443. doi: 10.1002/jev2.12443. J Extracell Vesicles. 2024. PMID: 38695388 Free PMC article. No abstract available.
Abstract
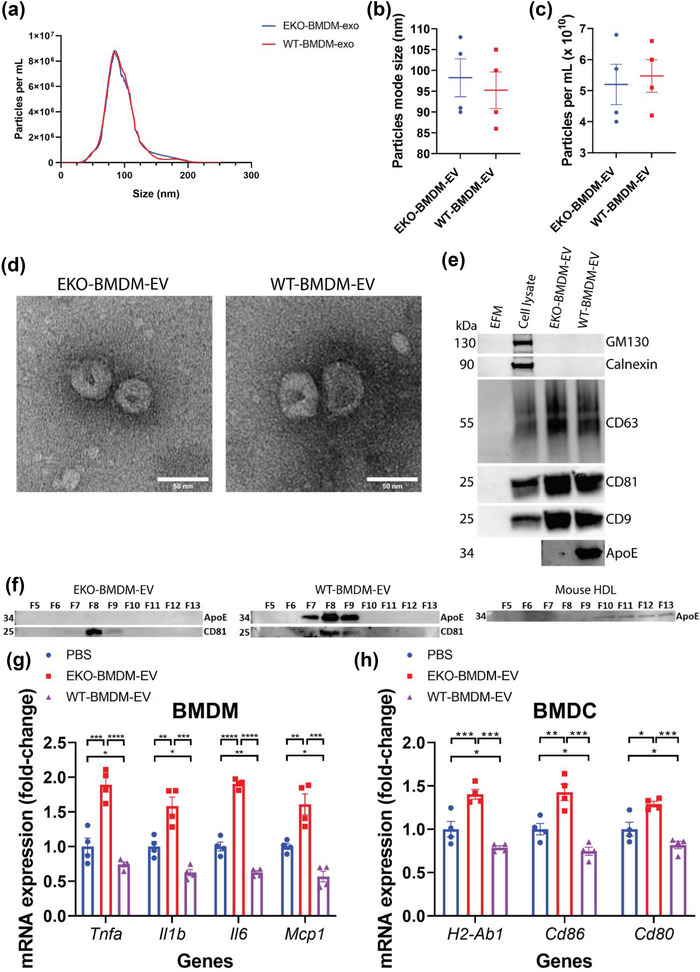
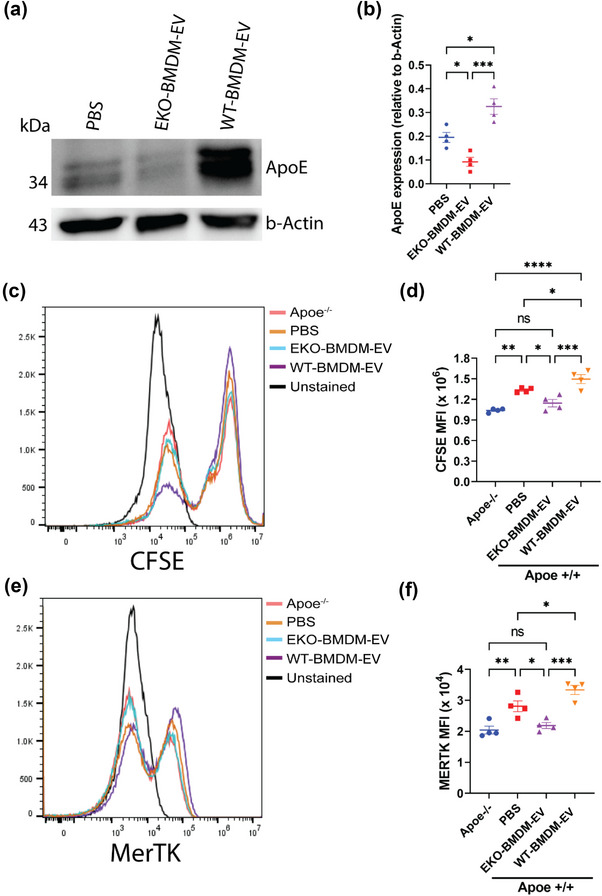
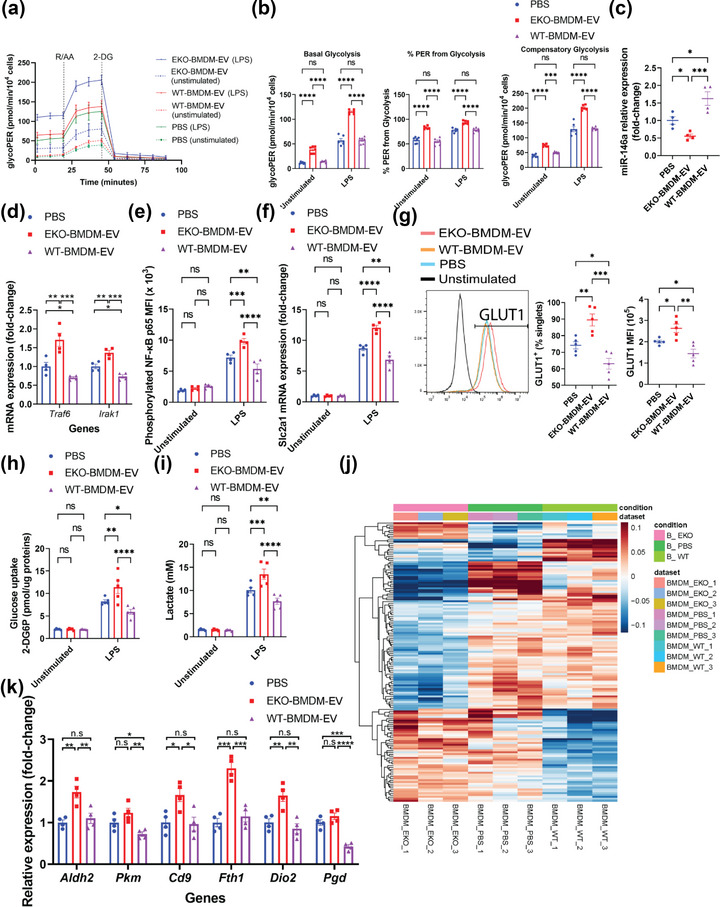
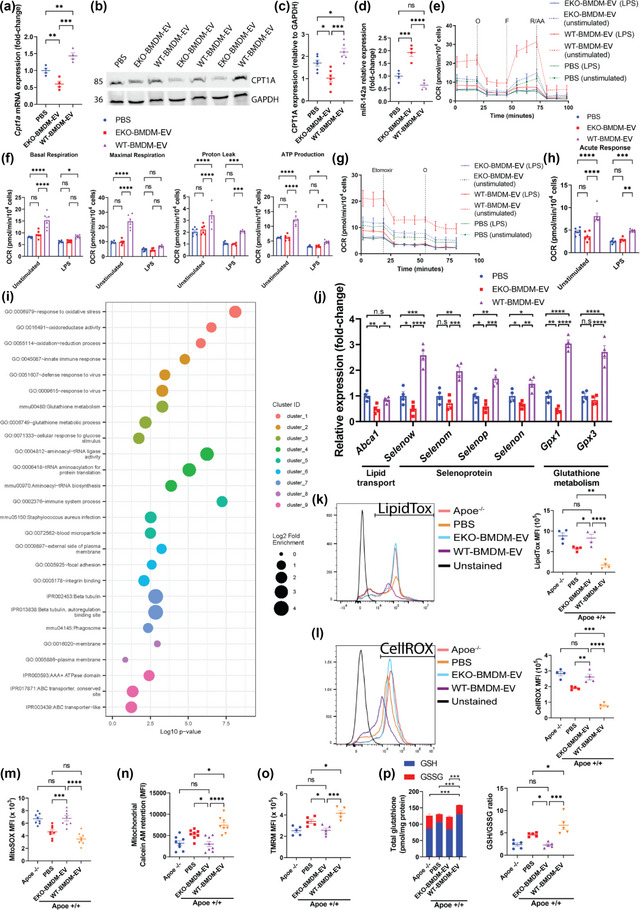
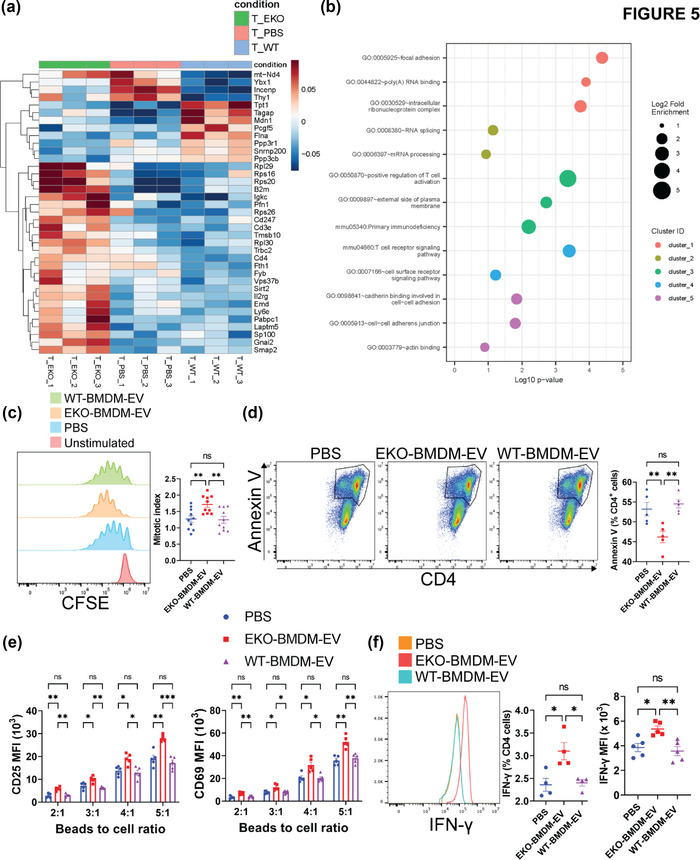
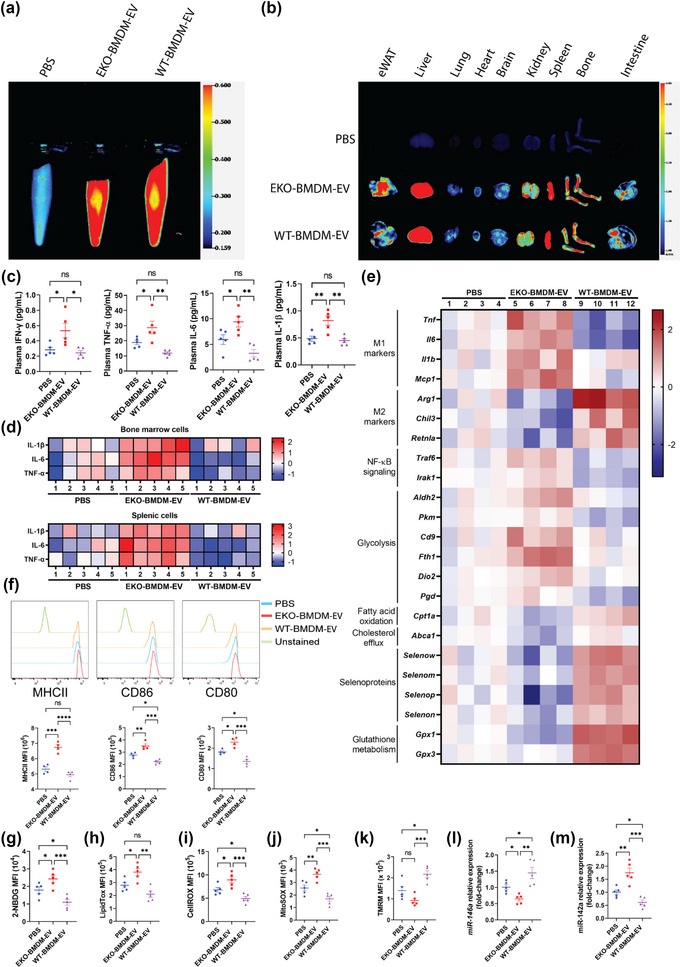
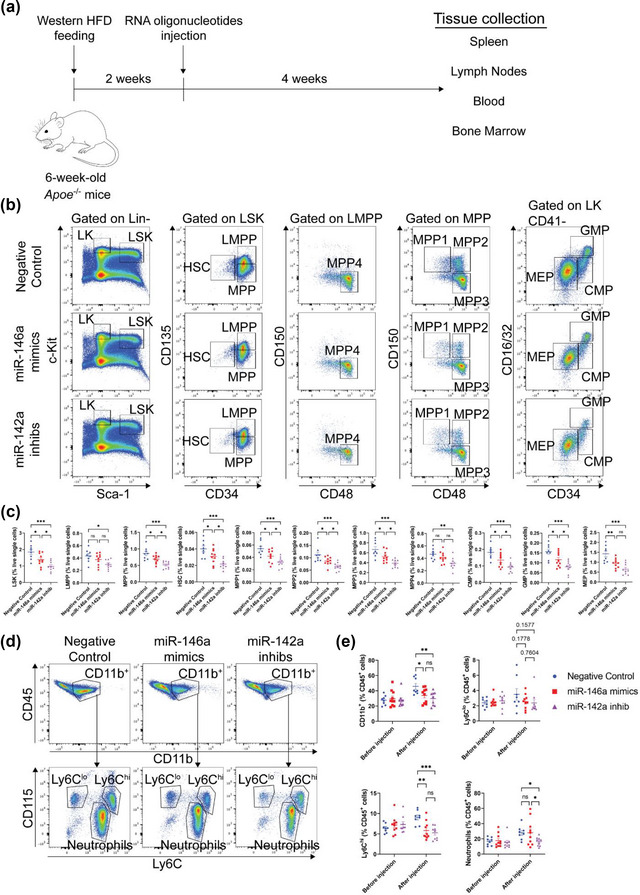
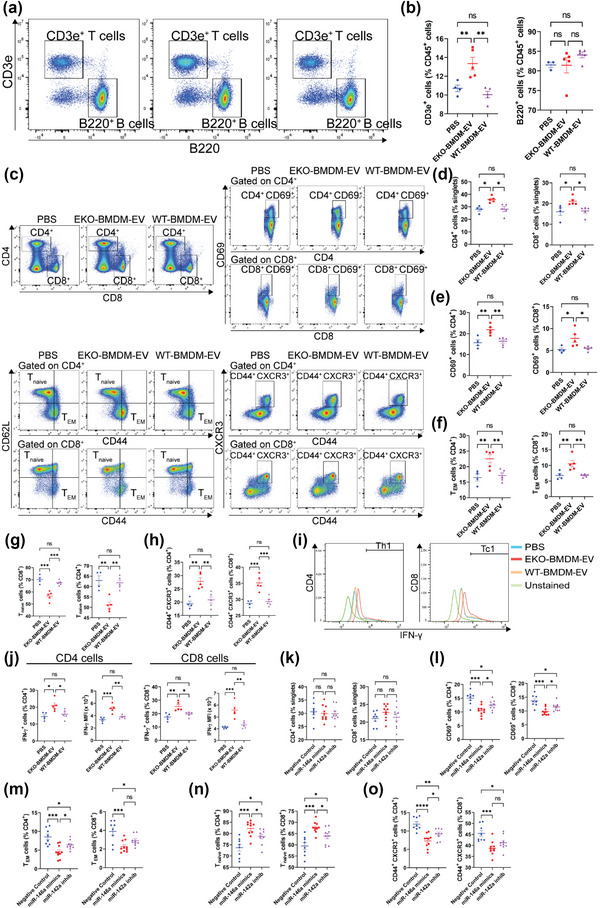
While apolipoprotein E (apoE) expression by myeloid cells is recognized to control inflammation, whether such benefits can be communicated via extracellular vesicles is not known. Through the study of extracellular vesicles produced by macrophages derived from the bone marrow of Wildtype (WT-BMDM-EV) and ApoE deficient (EKO-BMDM-EV) mice, we uncovered a critical role for apoE expression in regulating their cell signaling properties. WT-BMDM-EV communicated anti-inflammatory properties to recipient myeloid cells by increasing cellular levels of apoE and miR-146a-5p, that reduced NF-κB signalling. They also downregulated cellular levels of miR-142a-3p, resulting in increased levels of its target carnitine palmitoyl transferase 1A (CPT1A) which improved fatty acid oxidation (FAO) and oxidative phosphorylation (OxPHOS) in recipient cells. Such favorable metabolic polarization enhanced cell-surface MerTK levels and the phagocytic uptake of apoptotic cells. In contrast, EKO-BMDM-EV exerted opposite effects by reducing cellular levels of apoE and miR-146a-5p, which increased NF-κB-driven GLUT1-mediated glucose uptake, aerobic glycolysis, and oxidative stress. Furthermore, EKO-BMDM-EV increased cellular miR-142a-3p levels, which reduced CPT1A levels and impaired FAO and OxPHOS in recipient myeloid cells. When cultured with naïve CD4+ T lymphocytes, EKO-BMDM-EV drove their activation and proliferation, and fostered their transition to a Th1 phenotype. While infusions of WT-BMDM-EV into hyperlipidemic mice resolved inflammation, infusions of EKO-BMDM-EV increased hematopoiesis and drove inflammatory responses in myeloid cells and T lymphocytes. ApoE-dependent immunometabolic signaling by macrophage extracellular vesicles was dependent on transcriptional axes controlled by miR-146a-5p and miR-142a-3p that could be reproduced by infusing miR-146a mimics & miR-142a antagonists into hyperlipidemic apoE-deficient mice. Together, our findings unveil a novel property for apoE expression in macrophages that modulates the immunometabolic regulatory properties of their secreted extracellular vesicles.
Keywords: ApoE; extracellular vesicles; immunometabolism; inflammation; macrophage; microRNA; oxidative stress.
© 2023 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles. This article has been contributed to by U.S. Government employees and their work is in the public domain in the USA.
Conflict of interest statement
There is no conflict of interest in writing this manuscript.
Figures

References
-
- Akiyama, T. E. , Sakai, S. , Lambert, G. , Nicol, C. J. , Matsusue, K. , Pimprale, S. , Lee, Y.‐H. , Ricote, M. , Glass, C. K. , Brewer, H. B , & Gonzalez, F. J., Jr (2002). Conditional disruption of the peroxisome proliferator‐activated receptor gamma gene in mice results in lowered expression of ABCA1, ABCG1, and apoE in macrophages and reduced cholesterol efflux. Molecular and Cellular Biology, 22(8), 2607–2619. - PMC - PubMed
-
- Baitsch, D. , Bock, H. H. , Engel, T. , Telgmann, R. , Müller‐Tidow, C. , Varga, G. , Bot, M. , Herz, J. , Robenek, H. , Von Eckardstein, A. , & Nofer, J.‐R. (2011). Apolipoprotein E induces antiinflammatory phenotype in macrophages. Arteriosclerosis, Thrombosis, and Vascular Biology, 31(5), 1160–1168. - PMC - PubMed
-
- Bjørklund, M. M. , Hollensen, A. K. , Hagensen, M. K. , Dagnæs‐Hansen, F. , Christoffersen, C. , Mikkelsen, J. G. , & Bentzon, J. F. (2014). Induction of atherosclerosis in mice and hamsters without germline genetic engineering. Circulation Research, 114(11), 1684–1689. - PubMed
-
- Boldin, M. P. , Taganov, K. D. , Rao, D. S. , Yang, L. , Zhao, J. L. , Kalwani, M. , Garcia‐Flores, Y. , Luong, M. , Devrekanli, A. , Xu, J. , Sun, G. , Tay, J. , Linsley, P. S. , & Baltimore, D. (2011). miR‐146a is a significant brake on autoimmunity, myeloproliferation, and cancer in mice. Journal of Experimental Medicine, 208(6), 1189–1201. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
