

Neuropilin 2 in osteoblasts regulates trabecular bone mass in male mice
- PMID: 37600714
- PMCID: PMC10436209
- DOI: 10.3389/fendo.2023.1223021
Neuropilin 2 in osteoblasts regulates trabecular bone mass in male mice
Abstract
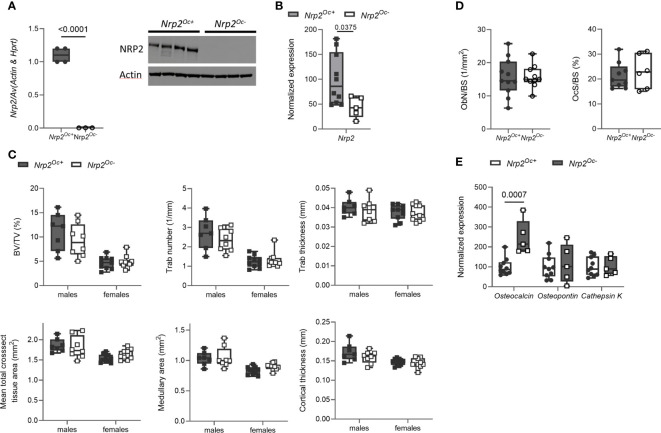
Introduction: Neuropilin 2 (NRP2) mediates the effects of class 3 semaphorins and vascular endothelial growth factor and is implicated in axonal guidance and angiogenesis. Moreover, NRP2 expression is suggested to be involved in the regulation of bone homeostasis. Indeed, osteoblasts and osteoclasts express NRP2 and male and female global Nrp2 knockout mice have a reduced bone mass accompanied by reduced osteoblast and increased osteoclast counts.
Methods: We first examined the in vitro effect of the calciotropic hormone 1,25-dihydroxyvitamin D3 [1,25(OH)2D3] on Nrp2 transcription in osteoblasts. We next generated mice with a conditional deletion of Nrp2 in the osteoblast cell lineage under control of the paired related homeobox 1 promoter and mice with a conditional Nrp2 knockdown in osteoclasts under control of the Lysozyme promoter. Mice were examined under basal conditions or after treatment with either the bone anabolic vitamin D3 analog WY 1048 or with 1,25(OH)2D3.
Results and discussion: We show that Nrp2 expression is induced by 1,25(OH)2D3 in osteoblasts and is associated with enrichment of the vitamin D receptor in an intronic region of the Nrp2 gene. In male mice, conditional deletion of Nrp2 in osteoblast precursors and mature osteoblasts recapitulated the bone phenotype of global Nrp2 knockout mice, with a reduced cortical cross-sectional tissue area and lower trabecular bone content. However, female mice with reduced osteoblastic Nrp2 expression display a reduced cross-sectional tissue area but have a normal trabecular bone mass. Treatment with the vitamin D3 analog WY 1048 (0.4 μg/kg/d, 14 days, ip) resulted in a similar increase in bone mass in both genotypes and genders. Deleting Nrp2 from the osteoclast lineage did not result in a bone phenotype, even though in vitro osteoclastogenesis of hematopoietic cells derived from mutant mice was significantly increased. Moreover, treatment with a high dose of 1,25(OH)2D3 (0.5 μg/kg/d, 6 days, ip), to induce osteoclast-mediated bone resorption, resulted in a similar reduction in trabecular and cortical bone mass. In conclusion, osteoblastic Nrp2 expression is suggested to regulate bone homeostasis in a sex-specific manner.
Keywords: bone; neuropilin 2; osteoblast; osteoclast; vitamin D.
Copyright © 2023 Verlinden, Doms, Janssens, Meyer, Pike, Carmeliet and Verstuyf.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






Similar articles
-
Nrp2 deficiency leads to trabecular bone loss and is accompanied by enhanced osteoclast and reduced osteoblast numbers.Bone. 2013 Aug;55(2):465-75. doi: 10.1016/j.bone.2013.03.023. Epub 2013 Apr 15. Bone. 2013. PMID: 23598046
-
The Vitamin D Receptor in Osteoblast-Lineage Cells Is Essential for the Proresorptive Activity of 1α,25(OH)2D3 In Vivo.Endocrinology. 2020 Nov 1;161(11):bqaa178. doi: 10.1210/endocr/bqaa178. Endocrinology. 2020. PMID: 32987399 Free PMC article.
-
Lack of the Thyroid Hormone Transporter Mct8 in Osteoblast and Osteoclast Progenitors Increases Trabecular Bone in Male Mice.Thyroid. 2020 Feb;30(2):329-342. doi: 10.1089/thy.2019.0271. Epub 2020 Jan 31. Thyroid. 2020. PMID: 31910109
-
Mechanism of inhibitory action of eldecalcitol, an active vitamin D analog, on bone resorption in vivo.J Steroid Biochem Mol Biol. 2013 Jul;136:171-4. doi: 10.1016/j.jsbmb.2012.11.010. Epub 2012 Dec 5. J Steroid Biochem Mol Biol. 2013. PMID: 23220095 Review.
-
1alpha(OH)D3 One-alpha-hydroxy-cholecalciferol--an active vitamin D analog. Clinical studies on prophylaxis and treatment of secondary hyperparathyroidism in uremic patients on chronic dialysis.Dan Med Bull. 2008 Nov;55(4):186-210. Dan Med Bull. 2008. PMID: 19232159 Review.
Cited by
-
MiR-331-3p facilitates osteoporosis and may promote osteoporotic fractures by modulating NRP2 expression.J Orthop Surg Res. 2024 Aug 17;19(1):487. doi: 10.1186/s13018-024-04959-7. J Orthop Surg Res. 2024. PMID: 39154011 Free PMC article.
-
Neuropilin2 in Mesenchymal Stromal Cells as a Potential Novel Therapeutic Target in Myelofibrosis.Cancers (Basel). 2024 May 18;16(10):1924. doi: 10.3390/cancers16101924. Cancers (Basel). 2024. PMID: 38792002 Free PMC article.
-
Relationship of the bone phenotype of the Klotho mutant mouse model of accelerated aging to changes in skeletal architecture that occur with chronological aging.Front Endocrinol (Lausanne). 2024 Jan 30;15:1310466. doi: 10.3389/fendo.2024.1310466. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 38352710 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous