

R-loops, type I topoisomerases and cancer
- PMID: 37600974
- PMCID: PMC9984992
- DOI: 10.1093/narcan/zcad013
R-loops, type I topoisomerases and cancer
Abstract
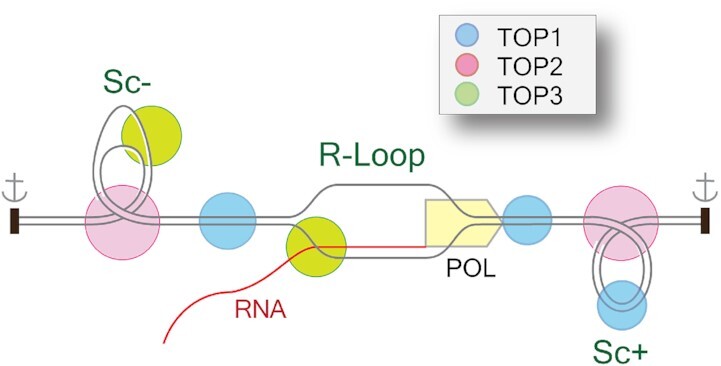
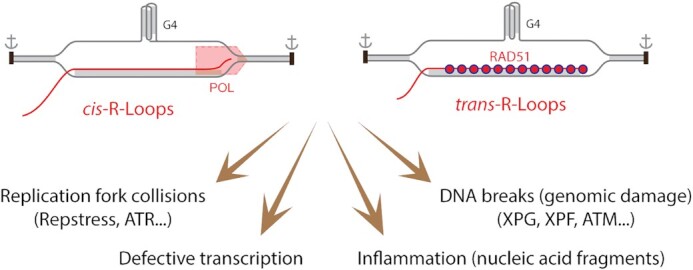
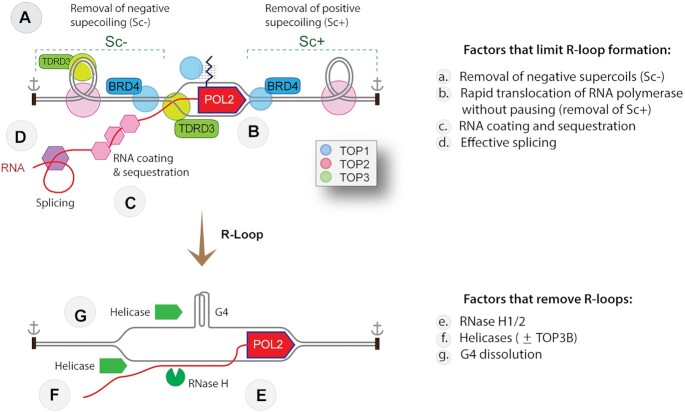
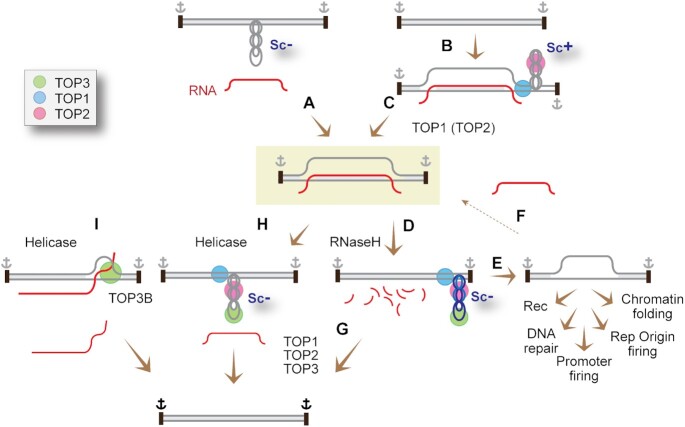
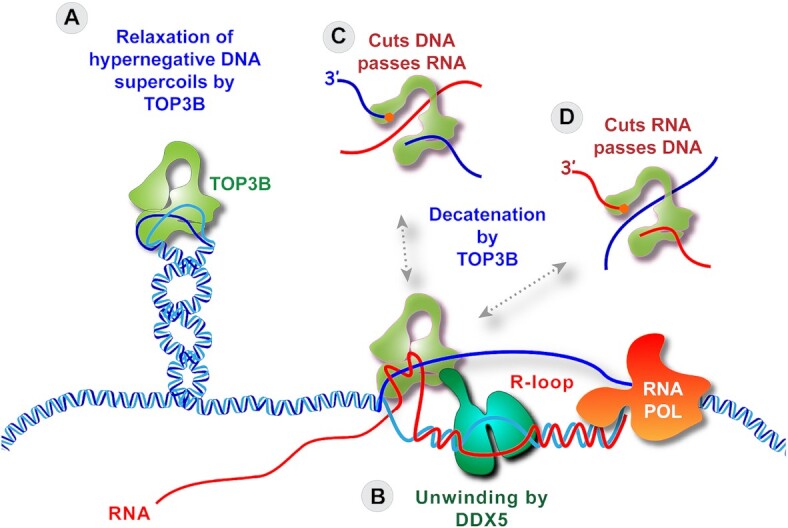
R-loops are abundant and dynamic structures ubiquitously present in human cells both in the nuclear and mitochondrial genomes. They form in cis in the wake of transcription complexes and in trans apart from transcription complexes. In this review, we focus on the relationship between R-loops and topoisomerases, and cancer genomics and therapies. We summarize the topological parameters associated with the formation and resolution of R-loops, which absorb and release high levels of genomic negative supercoiling (Sc-). We review the deleterious consequences of excessive R-loops and rationalize how human type IA (TOP3B) and type IB (TOP1) topoisomerases regulate and resolve R-loops in coordination with helicase and RNase H enzymes. We also review the drugs (topoisomerase inhibitors, splicing inhibitors, G4 stabilizing ligands) and cancer predisposing genes (BRCA1/2, transcription, and splicing genes) known to induce R-loops, and whether stabilizing R-loops and thereby inducing genomic damage can be viewed as a strategy for cancer treatment.
Published by Oxford University Press on behalf of NAR Cancer 2023.
Figures

References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
