

Fancm has dual roles in the limiting of meiotic crossovers and germ cell maintenance in mammals
- PMID: 37601968
- PMCID: PMC10435384
- DOI: 10.1016/j.xgen.2023.100349
Fancm has dual roles in the limiting of meiotic crossovers and germ cell maintenance in mammals
Abstract
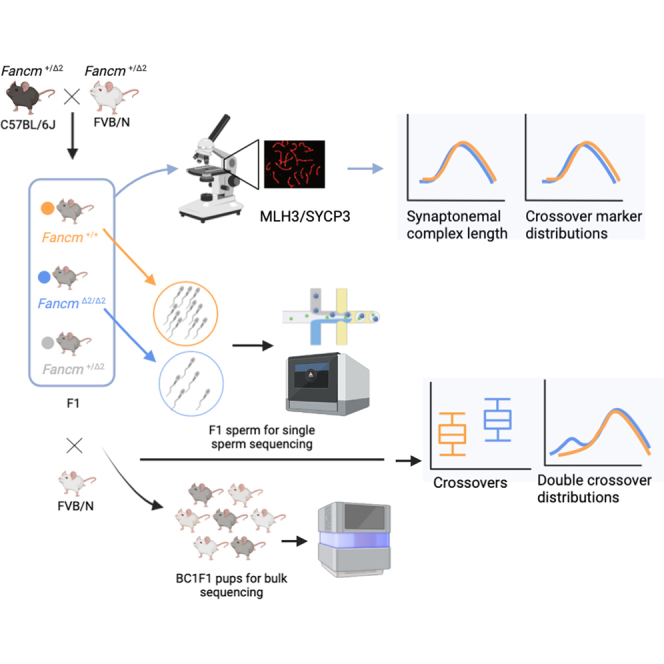
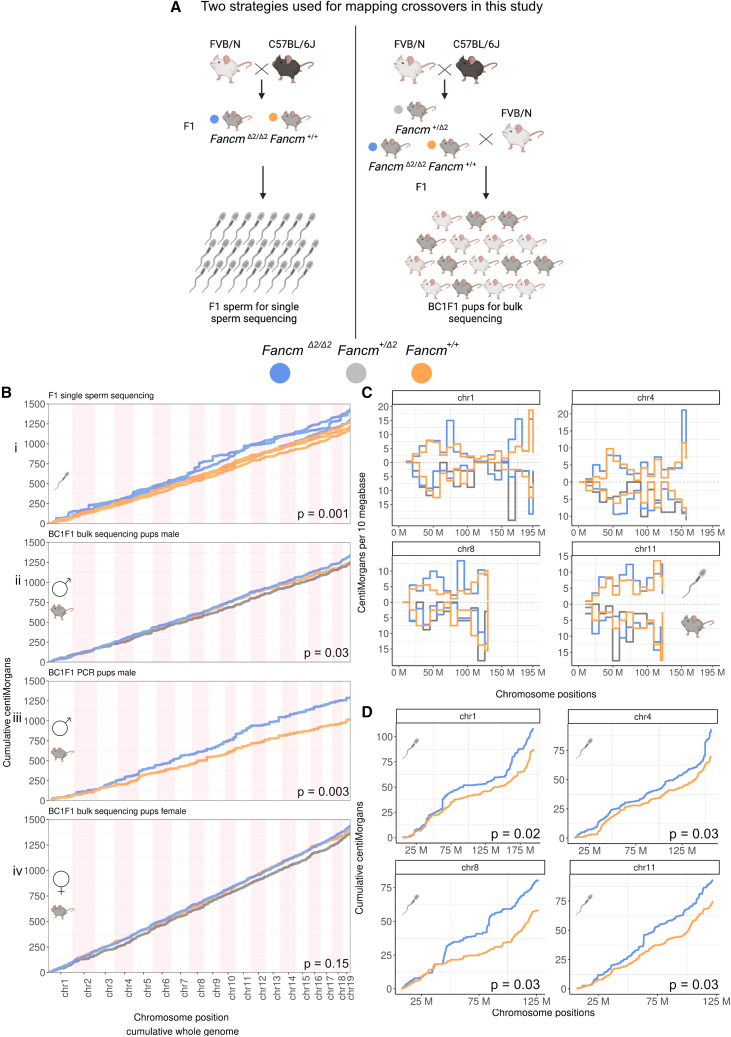
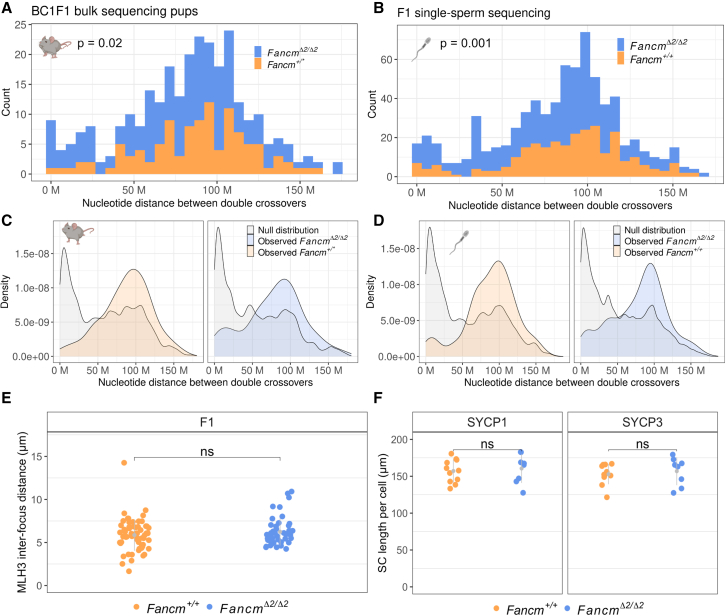
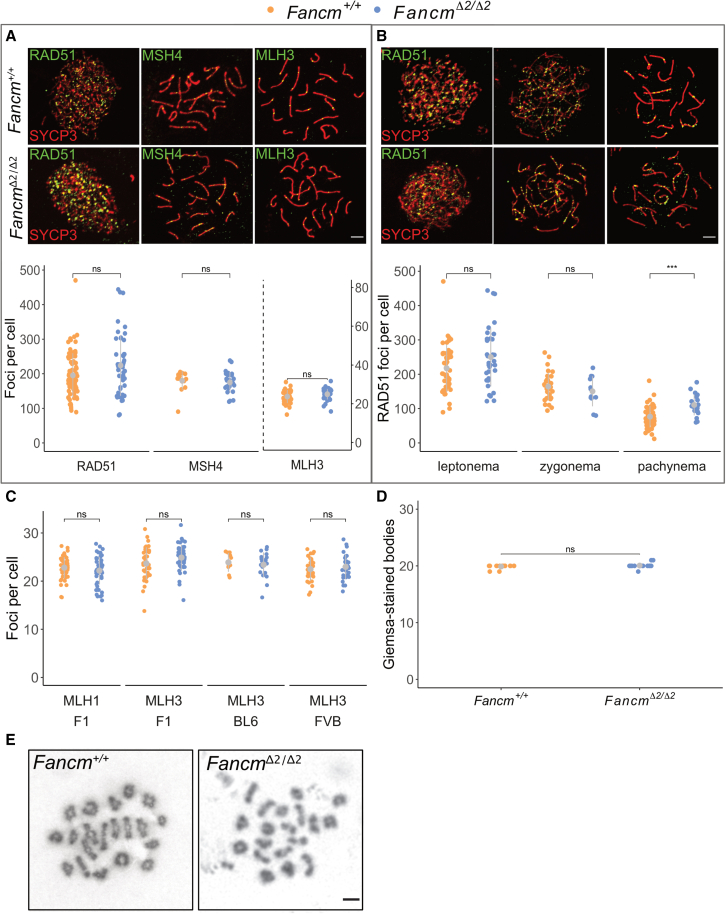
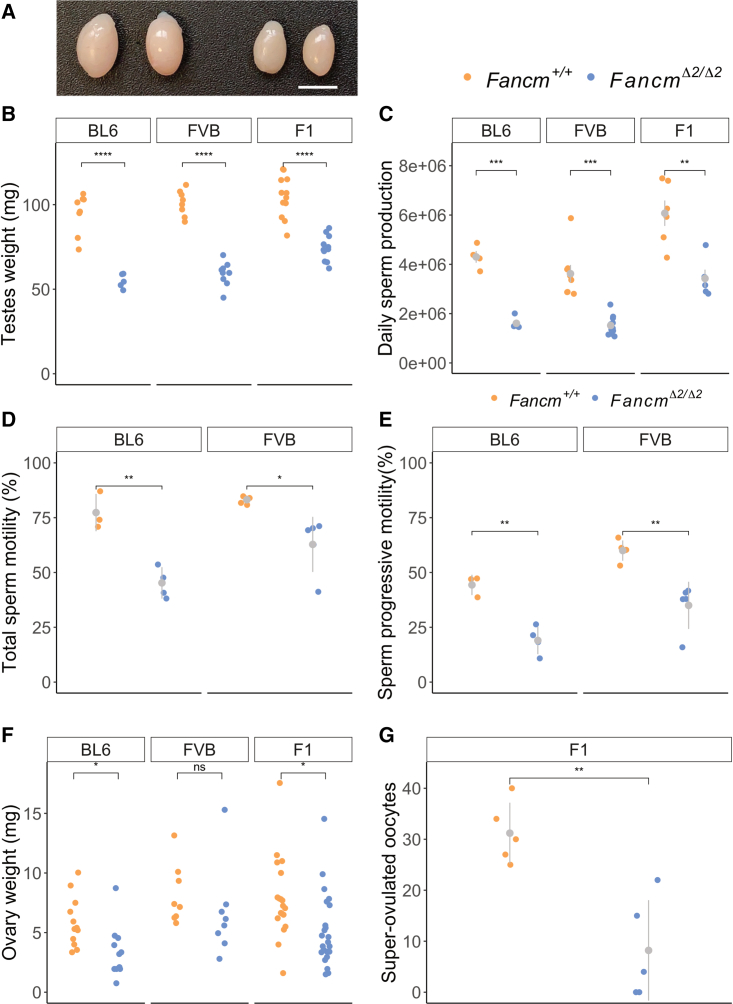
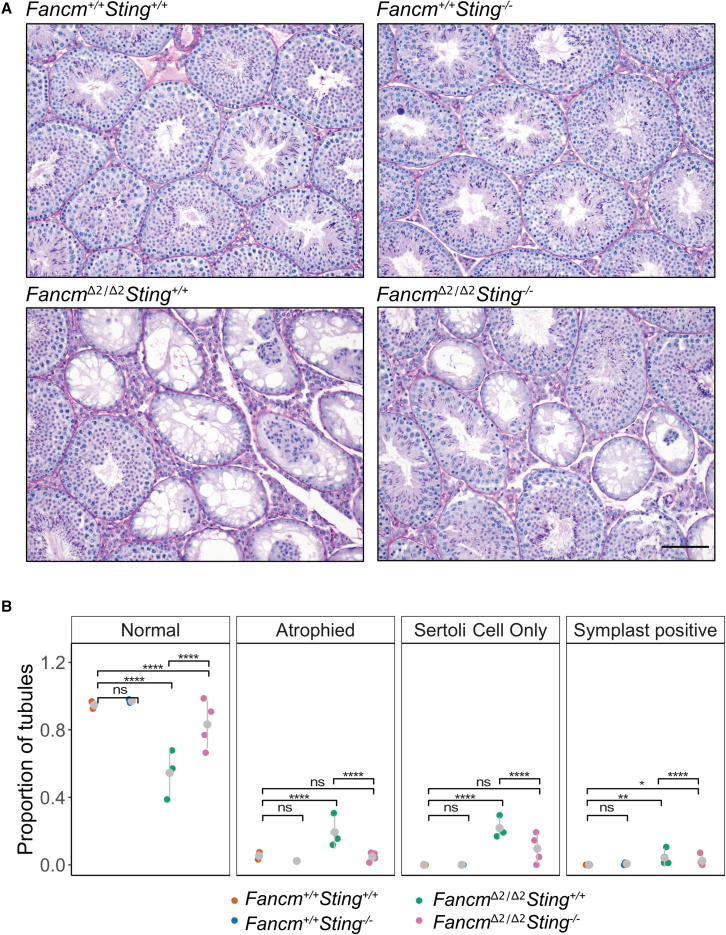
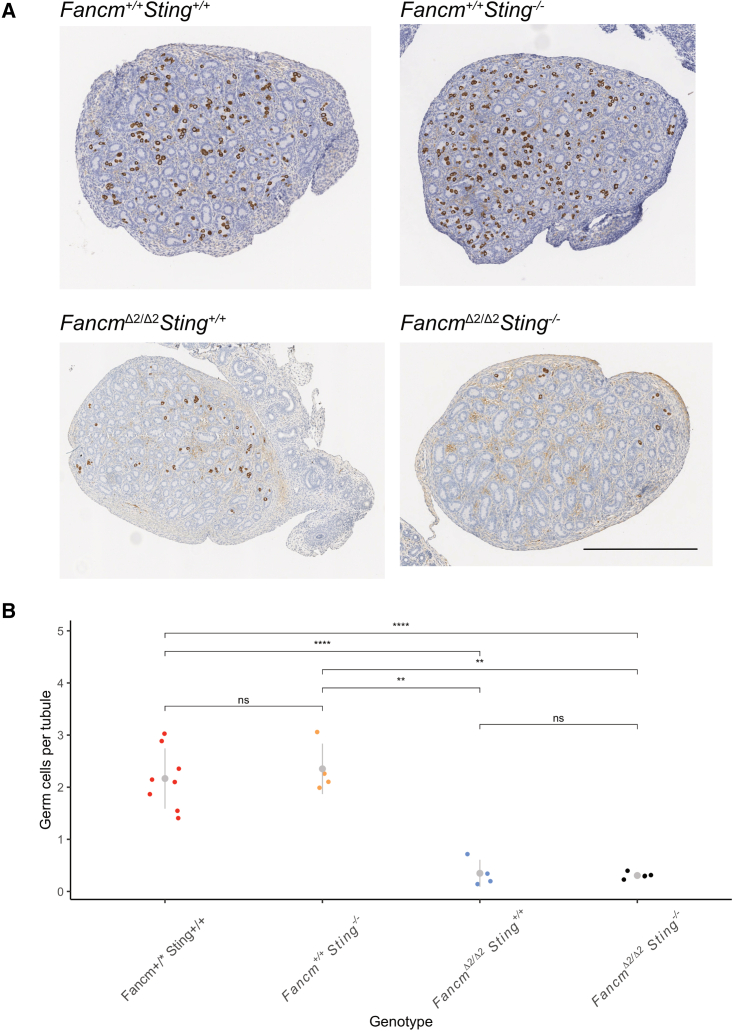
Meiotic crossovers are required for accurate chromosome segregation and producing new allelic combinations. Meiotic crossover numbers are tightly regulated within a narrow range, despite an excess of initiating DNA double-strand breaks. Here, we reveal the tumor suppressor FANCM as a meiotic anti-crossover factor in mammals. We use unique large-scale crossover analyses with both single-gamete sequencing and pedigree-based bulk-sequencing datasets to identify a genome-wide increase in crossover frequencies in Fancm-deficient mice. Gametogenesis is heavily perturbed in Fancm loss-of-function mice, which is consistent with the reproductive defects reported in humans with biallelic FANCM mutations. A portion of the gametogenesis defects can be attributed to the cGAS-STING pathway after birth. Despite the gametogenesis phenotypes in Fancm mutants, both sexes are capable of producing offspring. We propose that the anti-crossover function and role in gametogenesis of Fancm are separable and will inform diagnostic pathways for human genomic instability disorders.
© 2023 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
FANCM promotes class I interfering crossovers and suppresses class II non-interfering crossovers in wheat meiosis.Nat Commun. 2022 Jun 25;13(1):3644. doi: 10.1038/s41467-022-31438-6. Nat Commun. 2022. PMID: 35752733 Free PMC article.
-
Drosophila FANCM helicase prevents spontaneous mitotic crossovers generated by the MUS81 and SLX1 nucleases.Genetics. 2014 Nov;198(3):935-45. doi: 10.1534/genetics.114.168096. Epub 2014 Sep 8. Genetics. 2014. PMID: 25205745 Free PMC article.
-
FANCM Limits Meiotic Crossovers in Brassica Crops.Front Plant Sci. 2018 Mar 23;9:368. doi: 10.3389/fpls.2018.00368. eCollection 2018. Front Plant Sci. 2018. PMID: 29628933 Free PMC article.
-
The choice in meiosis - defining the factors that influence crossover or non-crossover formation.J Cell Sci. 2011 Feb 15;124(Pt 4):501-13. doi: 10.1242/jcs.074427. J Cell Sci. 2011. PMID: 21282472 Review.
-
Crossover patterns under meiotic chromosome program.Asian J Androl. 2021 Nov-Dec;23(6):562-571. doi: 10.4103/aja.aja_86_20. Asian J Androl. 2021. PMID: 33533735 Free PMC article. Review.
Cited by
-
The DNA helicase FANCJ (BRIP1) functions in double strand break repair processing, but not crossover formation during prophase I of meiosis in male mice.PLoS Genet. 2024 Feb 20;20(2):e1011175. doi: 10.1371/journal.pgen.1011175. eCollection 2024 Feb. PLoS Genet. 2024. PMID: 38377115 Free PMC article.
-
The DNA helicase FANCJ (BRIP1) functions in Double Strand Break repair processing, but not crossover formation during Prophase I of meiosis in male mice.bioRxiv [Preprint]. 2023 Oct 8:2023.10.06.561296. doi: 10.1101/2023.10.06.561296. bioRxiv. 2023. Update in: PLoS Genet. 2024 Feb 20;20(2):e1011175. doi: 10.1371/journal.pgen.1011175. PMID: 37873301 Free PMC article. Updated. Preprint.
-
FANCM Gene Variants in a Male Diagnosed with Sertoli Cell-Only Syndrome and Diffuse Astrocytoma.Genes (Basel). 2024 May 28;15(6):707. doi: 10.3390/genes15060707. Genes (Basel). 2024. PMID: 38927643 Free PMC article.
-
Structural basis of Fanconi anemia pathway activation by FANCM.EMBO J. 2025 Jul;44(14):4013-4036. doi: 10.1038/s44318-025-00468-3. Epub 2025 May 30. EMBO J. 2025. PMID: 40447800 Free PMC article.
-
[Homozygous Variant of FANCM of the Fanconi Anemia Pathway Causes Premature Ovarian Insufficiency: Investigation of the Pathogenic Mechanism].Sichuan Da Xue Xue Bao Yi Xue Ban. 2024 May 20;55(3):559-565. doi: 10.12182/20240560207. Sichuan Da Xue Xue Bao Yi Xue Ban. 2024. PMID: 38948269 Free PMC article. Chinese.
References
-
- Bakker S.T., van de Vrugt H.J., Rooimans M.A., Oostra A.B., Steltenpool J., Delzenne-Goette E., van der Wal A., van der Valk M., Joenje H., te Riele H., de Winter J.P. Fancm-deficient mice reveal unique features of Fanconi anemia complementation group M. Hum. Mol. Genet. 2009;18:3484–3495. doi: 10.1093/hmg/ddp297. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
