

Changes in cell wall composition due to a pectin biosynthesis enzyme GAUT10 impact root growth
- PMID: 37606259
- PMCID: PMC10663140
- DOI: 10.1093/plphys/kiad465
Changes in cell wall composition due to a pectin biosynthesis enzyme GAUT10 impact root growth
Abstract
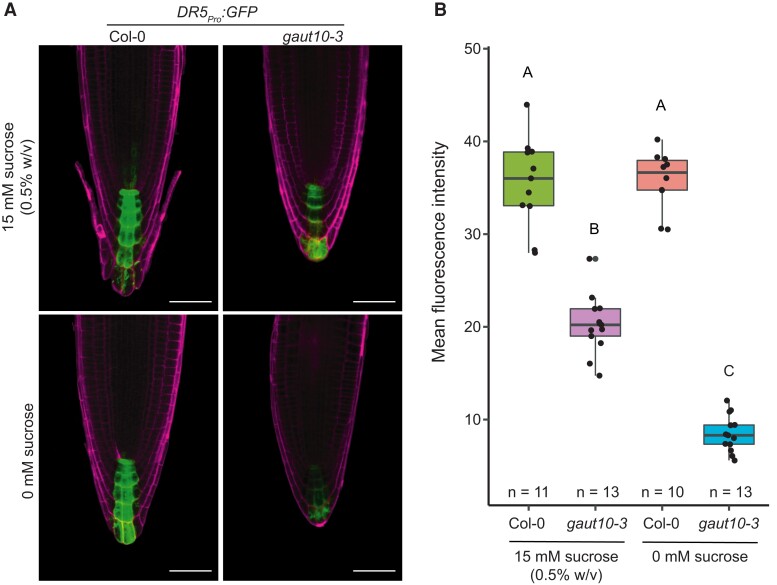
Arabidopsis (Arabidopsis thaliana) root development is regulated by multiple dynamic growth cues that require central metabolism pathways such as β-oxidation and auxin. Loss of the pectin biosynthesizing enzyme GALACTURONOSYLTRANSFERASE 10 (GAUT10) leads to a short-root phenotype under sucrose-limited conditions. The present study focused on determining the specific contributions of GAUT10 to pectin composition in primary roots and the underlying defects associated with gaut10 roots. Using live-cell microscopy, we determined reduced root growth in gaut10 is due to a reduction in both root apical meristem size and epidermal cell elongation. In addition, GAUT10 was required for normal pectin and hemicellulose composition in primary Arabidopsis roots. Specifically, loss of GAUT10 led to a reduction in galacturonic acid and xylose in root cell walls and altered the presence of rhamnogalacturonan-I (RG-I) and homogalacturonan (HG) polymers in the root. Transcriptomic analysis of gaut10 roots compared to wild type uncovered hundreds of genes differentially expressed in the mutant, including genes related to auxin metabolism and peroxisome function. Consistent with these results, both auxin signaling and metabolism were modified in gaut10 roots. The sucrose-dependent short-root phenotype in gaut10 was linked to β-oxidation based on hypersensitivity to indole-3-butyric acid (IBA) and an epistatic interaction with TRANSPORTER OF IBA1 (TOB1). Altogether, these data support a growing body of evidence suggesting that pectin composition may influence auxin pathways and peroxisome activity.
© The Author(s) 2023. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures








References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
