

Chloroplast-related host proteins interact with NIb and NIa-Pro of soybeans mosaic virus and induce resistance in the susceptible cultivar
- PMID: 37607595
- PMCID: PMC10472001
- DOI: 10.1016/j.virusres.2023.199205
Chloroplast-related host proteins interact with NIb and NIa-Pro of soybeans mosaic virus and induce resistance in the susceptible cultivar
Abstract
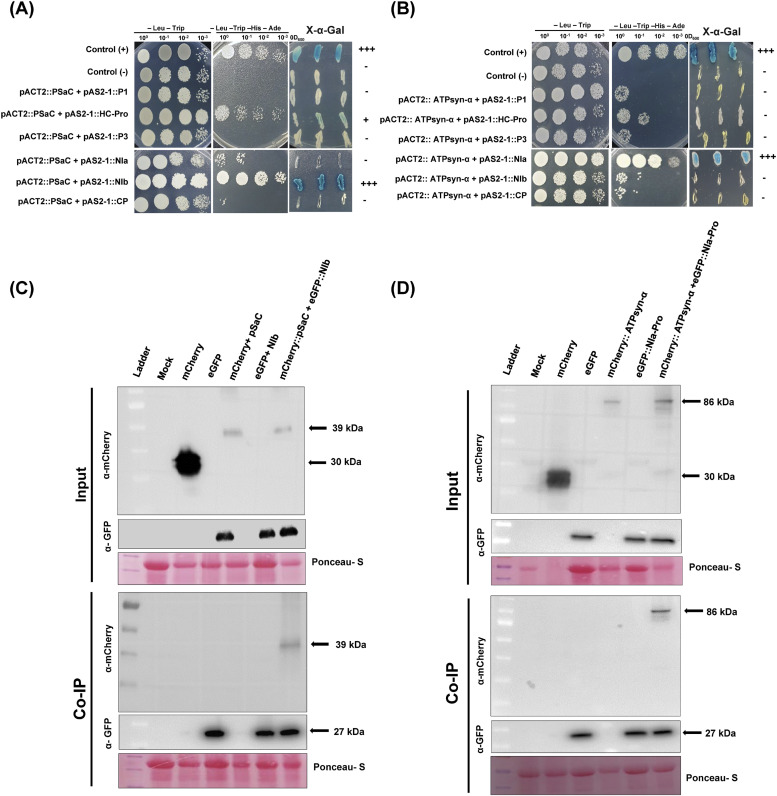
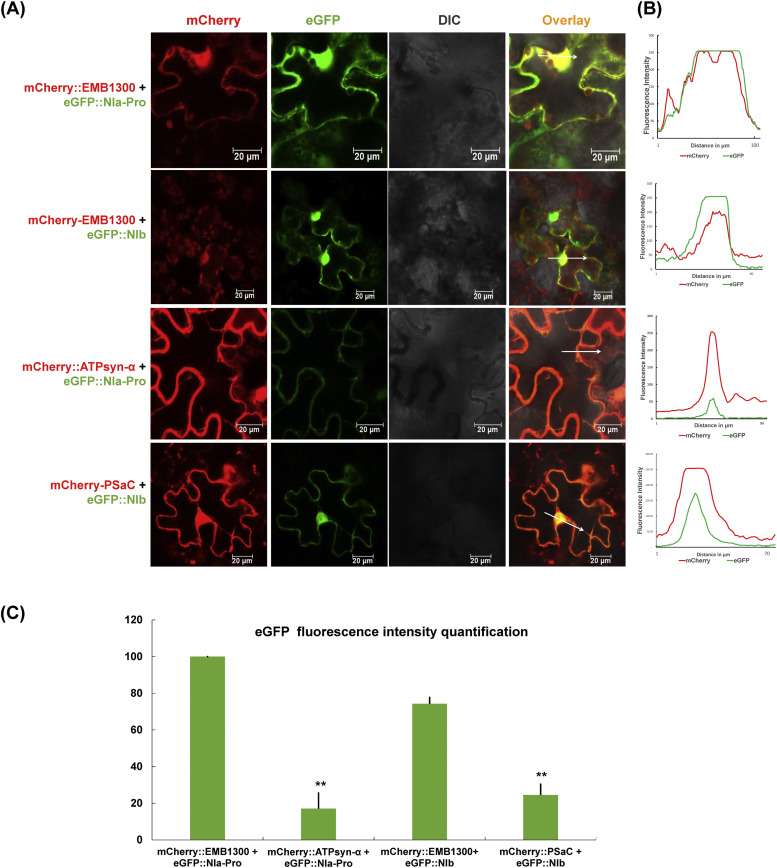
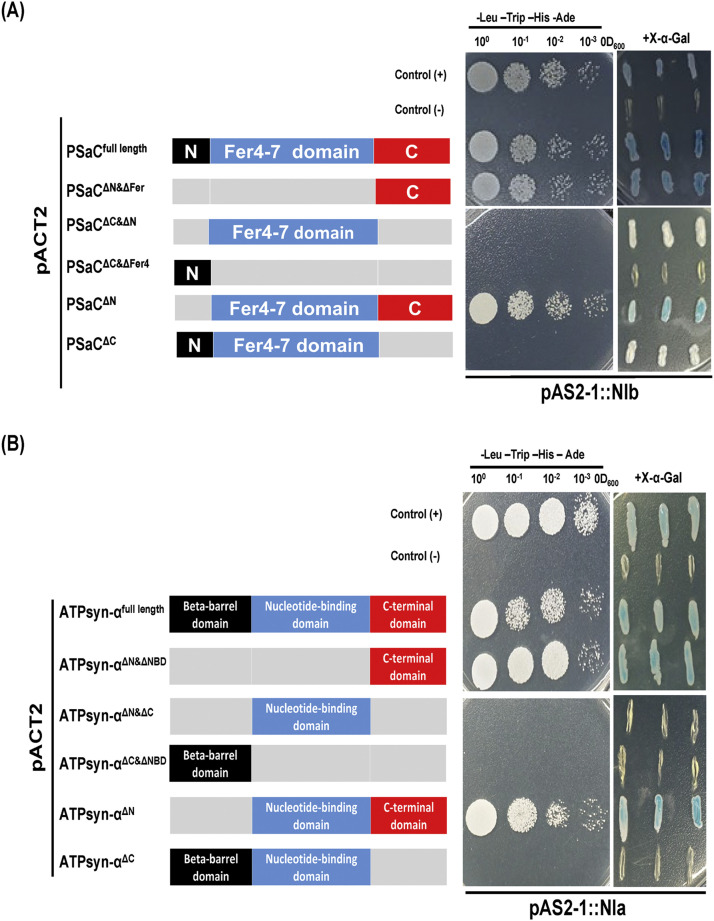
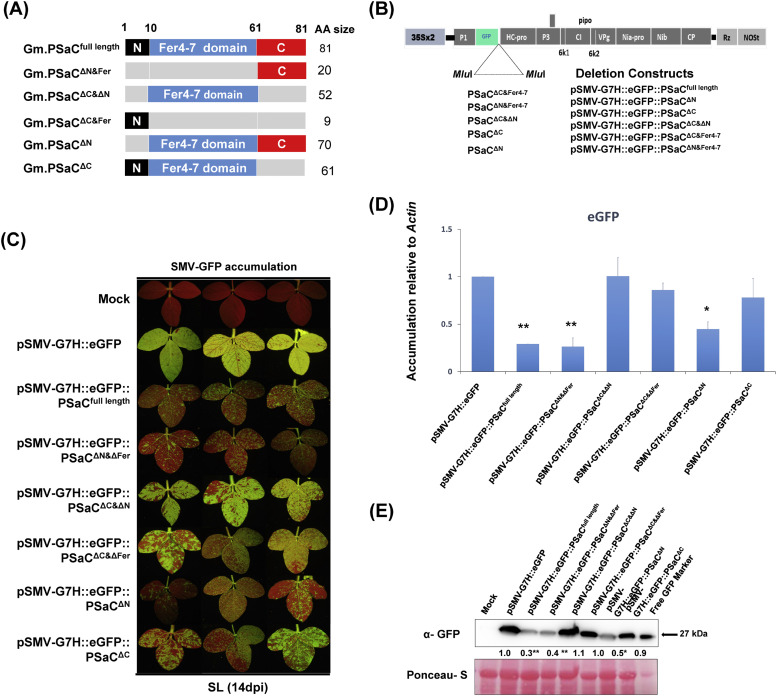
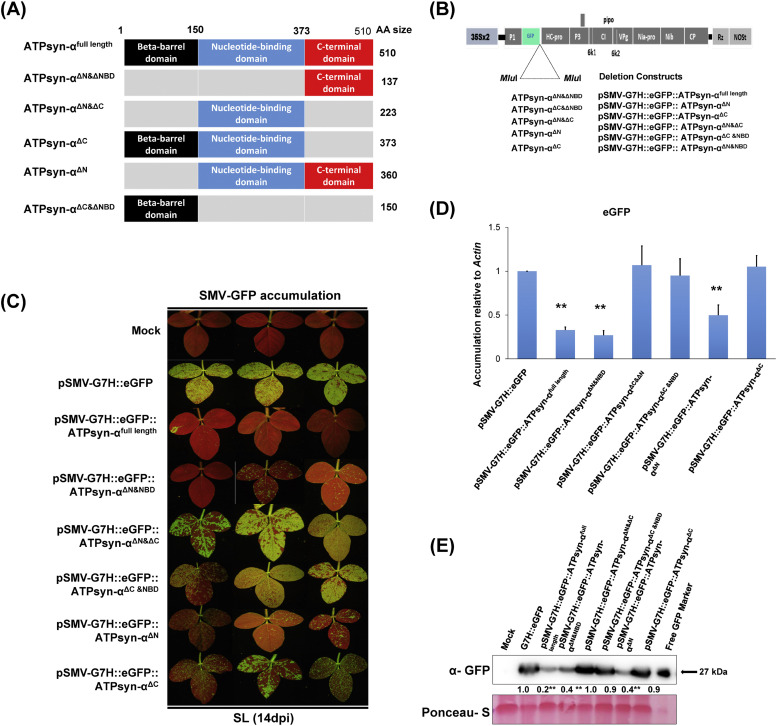
To gain a deeper understanding of the molecular mechanisms involved in viral infection and the corresponding plant resistance responses, it is essential to investigate the interactions between viral and host proteins. In the case of viral infections in plants, a significant portion of the affected gene products are closely associated with chloroplasts and photosynthesis. However, the molecular mechanisms underlying the interplay between the virus and host chloroplast proteins during replication remain poorly understood. In our previous study, we made an interesting discovery regarding soybean mosaic virus (SMV) infection in resistant and susceptible soybean cultivars. We found that the photosystem I (PSI) subunit (PSaC) and ATP synthase subunit α (ATPsyn-α) genes were up-regulated in the resistant cultivar following SMV-G7H and SMV-G5H infections compared to the susceptible cultivar. Overexpression of these two genes within the SMV-G7H genome in the susceptible cultivar Lee74 (rsv3-null) reduced SMV accumulation, whereas silencing of the PSaC and ATPsyn-α genes promoted SMV accumulation. We have also found that the PSaC and ATPsyn-α proteins are present in the chloroplast envelope, nucleus, and cytoplasm. Building on these findings, we now characterized protein-protein interactions between PSaC and ATPsyn-α with two viral proteins, NIb and NIa-Pro, respectively, of SMV. Through co-immunoprecipitation (Co-IP) experiments, we confirmed the interactions between these proteins. Moreover, when the C-terminal region of either PSaC or ATPsyn-α was overexpressed in the SMV-G7H genome, we observed a reduction in viral accumulation and systemic infection in the susceptible cultivar. Based on these results, we propose that the PSaC and ATPsyn-α genes play a modulatory role in conferring resistance to SMV infection by influencing the function of NIb and NIa-Pro-in SMV replication and movement. The identification of these photosynthesis-related genes as key players in the interplay between the virus and the host provides valuable insights for developing more targeted control strategies against SMV. Additionally, by utilizing these genes, it may be possible to genetically engineer plants with improved photosynthetic efficiency and enhanced resistance to SMV infection.
Keywords: Chloroplast-virus interplay; Plant defense; Soybean mosaic virus; Soybeans; Viral replication.
Copyright © 2023 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Similar articles
-
Photosynthesis-related genes induce resistance against soybean mosaic virus: Evidence for involvement of the RNA silencing pathway.Mol Plant Pathol. 2022 Apr;23(4):543-560. doi: 10.1111/mpp.13177. Epub 2021 Dec 28. Mol Plant Pathol. 2022. PMID: 34962034 Free PMC article.
-
Overexpression of purple acid phosphatase GmPAP2.1 confers resistance to Soybean mosaic virus in a susceptible soybean cultivar.J Exp Bot. 2022 Mar 2;73(5):1623-1642. doi: 10.1093/jxb/erab496. J Exp Bot. 2022. PMID: 34758072
-
Elements Involved in the Rsv3-Mediated Extreme Resistance against an Avirulent Strain of Soybean Mosaic Virus.Viruses. 2018 Oct 24;10(11):581. doi: 10.3390/v10110581. Viruses. 2018. PMID: 30355968 Free PMC article.
-
Decades of Genetic Research on Soybean mosaic virus Resistance in Soybean.Viruses. 2022 May 24;14(6):1122. doi: 10.3390/v14061122. Viruses. 2022. PMID: 35746594 Free PMC article. Review.
-
The Current Status of the Soybean-Soybean Mosaic Virus (SMV) Pathosystem.Front Microbiol. 2016 Nov 30;7:1906. doi: 10.3389/fmicb.2016.01906. eCollection 2016. Front Microbiol. 2016. PMID: 27965641 Free PMC article. Review.
References
-
- Broder Y.C., Katz S., Aronheim A. The ras recruitment system, a novel approach to the study of protein-protein interactions. Curr. Biol. 1998;8:1121–1124. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
